Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

4 м), а на коже спины — более 60 мм (6- Ю-2 м). Эти отличия связаны главным образом с различными размерами кожных рецептивных полей (от 0,5 мм2 до 3 см2) и со степенью их перекрытия. |
юлезнью движения (примером которой может служить морская болезнь). Последняя [роявляется изменением сердечного ритма (учащение, а затем замедление), сужением, : затем расширением сосудов, усилением движения желудка, головокружением, тошно- ой и рвотой. Повышенная склонность к болезни движения может быть уменьшена пециальной тренировкой (вращение, качели) и применением ряда лекарственных редств.
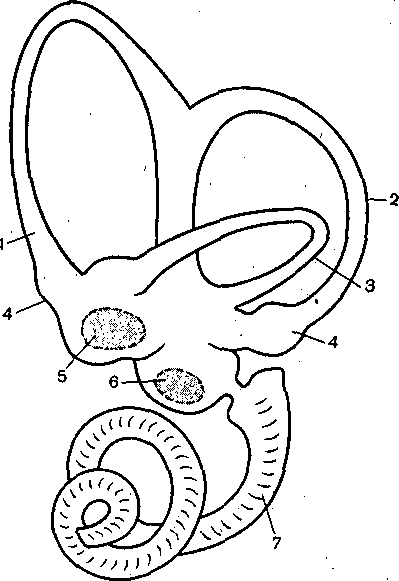
Вестибулоглаз.одвигательные рефлексы (глазной нистагм) состоят в ритмиче- ком медленном движении глаз в противоположную вращению сторону, сменяющемся качком глаз обратно. Само возникновение и характеристика вращательного глазного истагма — важные показатели, состояния вестибулярной системы и широко исполь- уются в авиационной, морской и космической медицине, а также в эксперименте и линике.
Интересно, что при переходе от земных условий, когда отолитовый аппарат посто- нно возбуждается ускорением силы тяжести, к условиям орбитальных полетов, харак- еризующихся невесомостью, деятельность отолитовых рецепторов изменяется: вначале до 70 ч) их активность резко снижается, а затем увеличивается и лишь к 5 суткам озвращается к норме (адаптация). Это было выявлено путем регистрации импульсов отдельных веточках вестибулярного нерва лягушек, запущенных в космос на специаль- ом биоспутнике.
В коре больших полушарий мозга обезьян основные афферентные проекции вестибулярного ппарата локализованы в задней части постцентральной извилины. В моторной коре кпереди г нижней части центральной борозды обнаружена вторая' вестибулярная зона. Исследование гих проекций осуществляется экспериментально путем электрического раздражения вестибуляр- )го нерва и регистрации вызванных потенциалов в разных отделах мозга. Выяснено, что есть )а основных пути поступления вестибулярных сигналов в кору обезьян: прямой — через дорсо- едиальную часть вентрального постлатерального ядра (VPL) и непрямой вестибуло-церебело- 1ламический путь через медиальную часть вентролатерального ядра (VL).
Локализация вестибулярной зоны коры человека окончательно не выяснена.
Вестибулярный анализатор помогает организму ориентироваться в пространстве эи активном движении животного и при пассивном переносе с места на место с завязан- э1ми глазами. При этом лабиринтный аппарат с помощью корковых отделов системы «ализирует и запоминает направление движения, повороты и пройденное расстояние, ледует подчеркнуть, что в нормальных условиях пространственная ориентировка зеспечивается совместной деятельностью зрительной и вестибулярной сенсорных щтем.
Чувствительность вестибулярного анализатора здорового человека весьма высока: отолито- >[й аппарат позволяет воспринять ускорение прямолинейного движения, равное всего 2 см/с2, орог различения наклона головы в сторону — всего около 1°, а вперед и назад — 1,5—2°. Рецеп- фная система полукружных каналов позволяет человеку замечать ускорения вращения в 2— в 1 с2.
СОМАТОСЕНСОРНЫЙ АНАЛИЗАТОР
В соматосенсорную анализаторную систему включают систему кожной чувстви- льности и чувствительную систему скелетно-мышечного аппарата, главная роль которой принадлежит проприорецепции. . -
Кожная рецепция
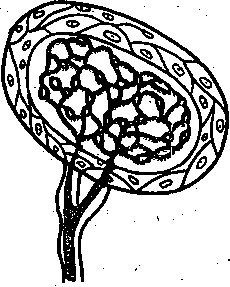
Кожные рецепторы. Рецепторная поверхность кожной чувствительной системы ■ромна — от 1,4 до 2,1 м2. В коже сосредоточено большое количество чувствительных прикосновению, давлению, вибрации, теплу и холоду, а также к болевым раздражениям фвных окончаний. Они весьма различны по структуре (рис. 228), локализуются на
Температурная рецепция. Температура тела человека характеризуется значительным постоянством и поэтому информация о температуре внешней среды, необходимая для деятельности механизмов терморегуляции, имеет особо важное значение. Терморецепторы располагаются в коже, на роговице глаза, в слизистых оболочках, а также в ЦНС — в, гипоталамусе. Они делятся на два вида: тепловые (их намного меньше и в коже они лежат глубже) и холодовые. Больше всего терморецепторов в коже лица и шеи. Гистологически тип терморецепторов до конца не выяснен, полагают, что ими могут быть немиелинизированные окончания дендритов афферентных нейронов.
Терморецепторы можно разделить на специфические и неспецифические. Первые возбуждаются лишь температурными воздействиями, вторые отвечают и на механическое раздражение. Большинство терморецепторов имеет локальные рецептивные поля и реагирует повышением частоты генерируемых импульсов, устойчиво длящимся все время действия стимула. Повышение частоты импульсации происходит пропорционально изменению температуры, причем постоянная частота у тепловых рецепторов наблюдается в диапазоне от 20 до 50 °С, а у холодовых — от 10 до 41 °С, Дифференциальная чувствительность терморецепторов велика: достаточно изменить температуру на 0,2 °С, чтобы вызвать длительные изменения их импульсации,
В определенных условиях холодовые пецепторы могут быть возбуждены и теплом (выше 45 °С), Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну.
В настоящее время считают, что наиболее важным фактором, определяющим активность терморецепторов, связанных 'С ними центральных структур и ощущения человека, является абсолютное значение температуры, а не ее изменения. В то же время интенсивность в начале температурных, ощущений зависит от разницы температуры кожи и температуры действующего на нее раздражителя, его площади и места приложения. Гак, в первый момент кажется холодной вода температуры 25 °С, если до этого руку держали в воде'температуры27 °С. Аналогичное ощущение возникает в первый момеь при переносе руки из воды; нагретой До 34 °С в воду температуры 31 6С. Однако у» через несколько секунд становится возможной истинная оценка абсолютной тег пературы. .
Болевая рецепция
Болевая, или ноцицептивная, чувствительность имеет особое значение для выж1 вания организма, так как сигнализирует об опасности при действии любых чрезмер* сильных и вредных агентов. В симптомокомплексе многих заболеваний боль — одь из первых, а иногда и единственное проявление патологии и важный индикатор в диа ностике. Однако корреляция между степенью болевых ощущений и тяжестью патолог чеСкого процесса отмечается не всегда. Несмотря на интенсивные исследования, до о пор не удается решить вопрос о существовании специфических болевых рецепторе и адекватных им болевых раздражителей.
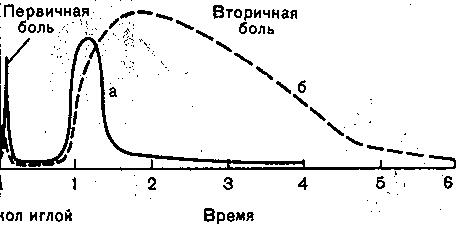
Сфсфмулированы две гипотезы об организации, болевого восприятия: 1) сущес вуют специфические болевые рецепторы (свободные нервные окончания с высоки порогом реакции); 2) специфических болевых рецепторов не существует и боль возн кает при сверхсильном раздражении любых рецепторов. В электрофизиологическ! Опытах на одиночных нервных волокнах -группы С обнаружено, что некоторые из hi реагируют преимущественно на чрезмерные механические, а другие — на чрезмерна тепловые действия. При болевых раздражениях небольшие по амплитуде импульс возникают также в волокнах группы "А. Соответственно разной скорости проведен! импульсов в волокнах групп С и А отмечается двойное ощущение боли: вначале четю по локализации и короткое, а затем — более длительное, разлитое и сильное (жгуче чувство боли (рис. 229). ' .
Механизм возбуждения рецепторов при болевых, воздействиях пока не выясне Предполагают, что особенно значимыми являются изменения рН ткани в области нер ного окончания, так как этот фактор обладает болевым эффектом при концентрат Н+-ионов, встречающихся в реальных условиях. Таким образом, наиболее общей прич ной возникновения боли можно считать изменение концентрации Н+-ионов при токе ческом воздействии на дыхательные ферменты или при механическом или термическе повреждении клеточных мембран. Не исключено, что одной из причин длительной жгуч1 боли может быть выделение при повреждении клеток гистамина, протеолитическ] ферментов, воздействующих на глобулины межклеточной жидкости и приводящ; к образованию ряда полипептидов (например, брадикинина), которые возбужда! окончания волокон группы С.
Адаптация болевых рецепторов возможна: ощущение укола от продолжают оставаться в коже иглы быстро проходит. Однако важной особенностью болевых реце торов во многих случаях является отсутствие существенной адаптации, что дела страдания больного особенно мучительными и требует применения анальгетиков.
Болевые раздражения вызывают ряд рефлекторных соматических и вегетативн] реакций, которые при умеренной их выраженности имеют приспособительное значен! но могут привести к вторичным грозным патологическим эффектам, например к шо! Отмечают повышение мышечного тонуса, частоты сердечного сокращения и дыхани повышение давления, сужение зрачков, увеличение содержания сахара в крови и р других эффектов.
При ноцицептивных воздействиях на кожу человек локализует их достаточно точь но при заболеваниях внутренних органов часты так называемые отражённые боли, прс цирующиеся в определенные части кожной поверхности ('зоны Захарьина — Геда). Та при стенокардии кроме болей в области сердца ^ощущается боль.в левой руке и лопат!
Наблюдаются и обратные эффекты. Так, при локальных тактильных, температур hi и болевых раздражениях определенных «активных» точек кожной поверхности вкл чаются цепи рефлекторных реакций, опосредуемых центральной и вегетативной нервн
Рис. 229. Развитие болевого ощущения во времени. ' ' - ■ '
а— нормальная кожа; б — кожа с повышенной болевой чувствительностью..
истемой. Они могут избирательно изменять кровоснабжение и трофику тех или иных р га нов и тканей. Методы и механизмы иглоукалывания (акупунктуры), локальных рижиганий и тонического массажа активных точек кожи в последние годы стали редметом глубокого научного исследования новой области медицины — рефлексо- ерапии.
Для уменьшения или устранения болевых ощущений в клинике используется мно- сество специальных веществ — анальгетиков, анестетиков и наркотиков. По ло.ка.лиза- ии действия они делятся на местные и общие. Первые (например, новокаин) блокируют озникновение и проведение болевых сигналов от рецепторов в спинной мозг или етрук- уры ствола мозга. Общие анестетики (например, эфир) снимают ощущение боли, локируя передачу импульсов между нейронами коры и ретикулярной формации мозга погружают человека в наркотический сон).
В последние годы выяснена высокая анал&гезирующая активность так называемых ейропептидов, большинство которых представляет собой либо гормоны (вазопрессин, кситоцин, АКТГ), либо их фрагменты. Часть нейропептидов является фрагментами -липотропного гормона (эндорфины). Анальгезирующее действие нейропептидов осно- ано на том, что они даже в минимальных дозах (в микрограммах) меняют эффектив- ость передачи в синапсах с «классическими» нейромедиаторами (ацетилхолин, нор- дреналин), в частности между первым и вторым сенсорными нейронами (задние столбы пинного мозга и другие структуры). С использованием нейропептидов в настоящее ремя связываются надежды на эффективное лечение ряда нервно-психических за- олеваний.
МЫШЕЧНАЯ И СУСТАВНАЯ РЕЦЕПЦИЯ (ПРОПРИОРЕЦЕПЦИЯ)
В мышцах млекопитающих животных и человека содержатся три типа специали- ированных рецепторов: первичные окончания веретен, вторичные окончания веретен сухожильные рецепторы Гольджи. Эти рецепторы реагируют на механические раздра- сения и участвуют в координации движений, являясь источником информации о состоя- ии двигательного аппарата.
В последние годы разработана методика отведения потенциалов действия отдельных фферентных волокон в не^ве человека, что позволило исследовать функцию веретен естественных условиях работы мышц.
Мышечные веретена представляют собой небольшие продолговатые образования длиной несколько миллиметров, шириной — десятые доли миллиметра), расположение в толще мышцы (рис. 230) . В разных скелетных мышцах число веретен на 1 г ткани арьирует от сотни до нескольких единиц. Внутри капсулы каждого веретена находится учок мышечных волокон. Эти волокна называют интрафузальными в отличие от всех стальных волокон мышцы, которые носят название экстрафузальных. Веретена расПо- ожены параллельно экстрафузальным волокнам, поэтому при растяжении мышцы агрузка на веретена увеличивается, а при сокращении — уменьшается.
Различают интрафузальные волокна двух типов: более толстые и длинные с ядрами, осредоточенными в средней, утолщенной .части волокна, ядерно-сумчатые и более ороткие и тонкие с ядрами, расположенными цепочкой,— ядерно-цепочечные. На
интрафузальных,волокнах спирально расположены чувствительные окончания афферентов группы 1а — так называемые первичные окончания, и чувствительные окончания афферентов группы II—так называемые вторичные окончания. Импульсация, идущая от веретен по афферентам группы 1а, на спинальном уровне моносинаптически и по- лисинаптически возбуждает мотонейроны своей мышцы и дисинаптически (через тор-, мозящий интернейрон) тормозит мотонейро- ны мышцы-антагониста (реципрокное торможение). Афференты группы II полисинап- тически возбуждают мотонейроны сгибателей и тормозят мотонейроны разгибателей. Имеются; однако, данные, что афференты в группе JI, идущие от разгибателей, могут возбуждать мотонейроны, своей мышцы. .
Веретена имеют и эфферентную иннервацию: интрафузальные мышечные волокна иннервируются аксонами, идущими к ним от у-мотонейронов. Эти так называемые у-эф- ференты подразделяются на динамические и статические.
В расслабленной мышце количество импульсов, идущих от веретен, невелико. Веретена реагируют импульсацией на удлинение (растяжение) мышцы, причем у первичных окончаний частота импульсации зависит главным образом от скорости удлинения, а у вторичных — от длины мышцы (динамический и статический ответы). Активация у-эфферентов приводит к повышению чувствительности веретен, причем динамические у-эфференты преимущественно усиливают реакцию на скорость удлинения мышцы, а статические— на длину.
И без растяжения мышЦы,' активация у-эфферентов сама по себе, вызывает им- пульсацию афферентов веретен'вследствие сокращения интрафузальных мышечных волокон. Показано, что возбуждение а-мотонейронов сопровождается возбуждением у-мотонейронов (а—у-коактивация). Уровеш возбуждения у-системы тем выше, чем интенсивнее возбуждены а-мотонейррны данной мышцы,, т. е. больше сила ее сокращения.
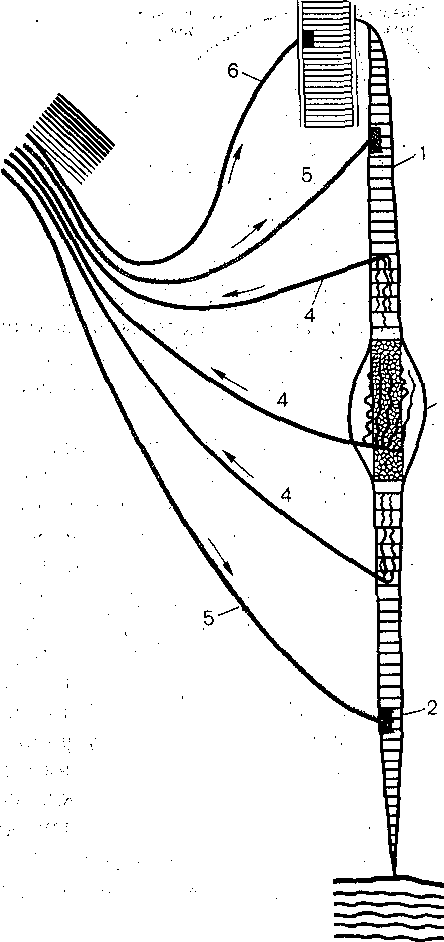
Рис. 230. Мышечное веретено.
Проксимальный конец интрафузального мышечного волокна (Г), прикрепленного к волокну скелетной мышцы; 2— Дистальный конец этого волокна прикрепленного . к фасции; 3 — ядерная сумка: 4 — афферентные волокна; 5 — гамма-эфферентные волокна; 6 — :моторное волокно, идущее к скелетной мышце.
Таким образом, веретена реагируют на два воздействия: периферическое— изменение длины мышцы и центральное — изменение уровня активации у-системы. Поэтому реакции веретен в естественных, условиях деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецепторов веретен, вызывающая рефлекс на растяжение. При активном Концентрическом сокращении мышцы (т. е. сокращении с укорочением) уменьшение длины,,мышцы оказывает на рецепторы веретена дезактивирующее действие, а возбуждение у-мотонейронов, сопутствующее возбуждению а-мотонейронов, вызывают активацию рецепторов. Поэтому импульсация от рецепторов веретен во время движения зависит от соотношения длины мышцы,
юрости ее укорочения и силы сокращения. Таким образом, веретена можно рассмотришь как непосредственный источник информации о длине мышцы и ее изменениях, если >лько мышца не возбуждена. При активном состоянии, мышцы необходимо учитывать 1ияние 7-системы: Согласно современным представлениям, во время активных движе- \\\ 7-мотонейроны поддерживают импульсацию веретен укорачивающейся мышцы, что эет возможность рецепторам реагировать на Вызванные помехами неравномерности зижения как увеличением, так и, уменьшением частоты импульсации и участвовать 1ким образом в коррекции движений (см. рис. 51 на с. 75).
Сухожильные рецепторы Гольджи находятся в зоне соединения мышечных волокон сухожилием и расположены последовательно по отношению к мышечным волокнам, ни слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении, нтенсивность их импульсации примерно пропорциональна силе сокращения мышцы, что ает основание рассматривать сухожильные рецепторы.как источник информации о силе, азвиваемой мышцей. Идущие от этих рецепторов афференты относятся к группе 16. [а спинальном.уровне они через интернейроны вызывают торможение мотонейронов эбственной мышцы и возбуждение мотонейронов антагониста.
Информация от мышечных рецепторов по восходящим путям спинного мозга оступает и в верхние отделы ЦНС, включая кору большого мозга, и участвует в ки- естезии.
Суставные рецепторы изучены меньше, чем мышечные. Известно, что они реагируют а положение сустава и на изменения суставного угла, участвуя таким образом в си- геме обратных связей от двигательного аппарата.
ПЕРЕРАБОТКА СОМАТОСЕНСОРНОЙ ИНФОРМАЦИИ
Чувствительность кожи и ощущение движения связаны с проведением^ в мозС игналов от рецепторов по двум Основным путям (трактам): лемнисковому и спйнотала- (ическому, значительно различающимся по своим морфологическим и функциональным войствам.
В последнее время описан и третий путь — латеральный тракт Морина, близкий по ряду арактеристик к лемнисковой системе.
Лемнисковый путь на всех уровнях состоит из относительно толстых и быстропро- одящих миелинизированных нервных волокон. Он передает в мозг сигналы о прикосно- ении к коже, давлении на нее и движениях в суставах. Отличительная особенность того пути — быстрая передача наиболее тонкой информации, дифференцированной о силе и месту воздействия. Первые нейроны этого пути находятся в спинальном ган- лии, их аксоны в составе задних столбов восходят к тонкому (Голля) и клиновидному Бурдаха) ядрам продолговатого мозга, где сигналы передаются на вторые нейроны емнискового пути. Часть волокон, в основном несущих сигналы от суставных рецепто- юв, оканчивается на мотонейронах сегментарного спинального уровня.
ПроприоЦептивная чувствительность передается в спинном, мозге также ,по дорсальному пиномозжечковому, спиноцервикзльному трактам и некоторым другим путям.
В продолговатом мозге в ядре тонкого пучка сосредоточены в основном Вторые 1ейроны тактильной чувствительности, а в клиновидном ядре — вторые нейроны про- фиоцептивной чувствительности: Аксоны этих нейронов образуют медиальный лемни- :ковый тракт («медиальную петлю») и после перекреста на уровне олив направляются. ( специфические ядра таламуса (вентробазальный ядерный комплекс — VP) . В этих щрах концентрируются третьи по порядку нейроны лемнискового пути. Их аксоны шправляются в соматосенсорную зону коры большого мозга. ,
По мере перехода на все более высокие уровни изменяются некоторые важные свойства нейронов лемнискового пути. Значительно увеличиваются рецептивные Поля гейронов (в продолговатом мозге в 2—30, а в коре большого мозга в 15—100 раз). Этветы клеток становятся все более продолжительными: даже короткое прикосновение
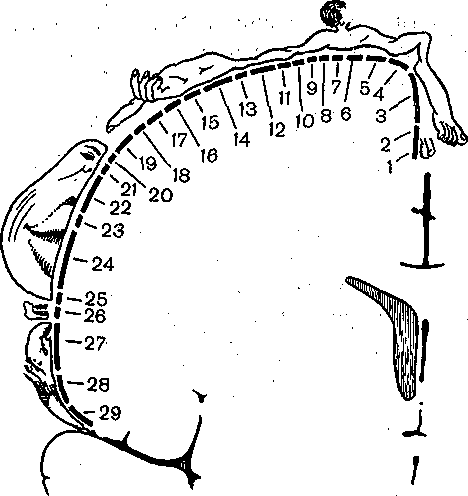
Рис. 231. Корковое представительство кожной чувствительности.
Расположение в соматосенсорной зоне коры больших полушарий человека проекций различных частей тела.
1 — половые органы; 2 — пальцы; 3 — ступня; 4 — голень; 5 — бедро; 6 — туловище; 7 — шея; 8 — голова; 9,— плечо; 10-11 — локоть; 12 — предплечье; 13,— запястье; 14 — кисть; 15—19 — пальцы; 20 — глаз; 21 — нос; 22 — лицо; 23 — верхняя губа; 24,26 — зубы; 25 — нижняя губа; 27 — язык; 28 — глотка; 29 — внутренние органы.. Размеры изображений частей тела соответствуют размерам сенсорного представительства.
к коже вызывает залп импульсов, длящийся несколько секунд. Отмечено появление так называемых нейронов новизны, реагирующих на смену раздражителя. Несмотря на увеличение размеров рецептивных полей, нейроны остаются достаточно специфичными (нейроны поверхностного прикосновения, глубокого прикосновения, нейроны движения в суставах и нейроны положения или угла сгибания суставов). Для корковой части лемнискового пути характерна четкая топографическая организация, т. е. проекция кожной поверхности осуществляется «точка в точку». При этом площадь коркового представительства той или иной части тела определяется ее функциональной значимостью (рис. 231).
Удаление соматосенсорной коры приводит к нарушению способности локализовать тактильные ощущения, а ее электростимуляция вызывает ощущение прикосновения, вибрации и зуда. В целом роль соматосенсорной коры состоит в интегральной оценке сложной афферентной соматосенсорной информации.
Спиноталамический путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинальном ганглии, откуда они посылают в спинной мозг медленно проводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спиноталамического тракта направляются после перекреста на спинальном уровне В вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спиноталамического пути лишь частично дают проекции в соматосенсорную зону коры.
Считается, что спиноталамический путь с более медленной передачей афферентных сигналов, со значительно менее четко дифференцируемой информацией о разных свойствах раздражителя и с менее четкой топографической локализацией служит для передачи температурной, всей болевой и в значительной мере — тактильной чувствительности. Болевая чувствительность практически не представлена на корковом уровне (раздражение коры не вызывает боли), поэтому считают, что высшим центром болевой чувствительности является таламус, где 60 % нейронов в соответствующих ядрах четко реагирует на болевое раздражение. Таким образом, эта система играет важную роль в организации генерализованных ответов на действие болевых, температурных и тактильных р!аздражителей, сигналы о которых идут через структуры ствола, подкорковые образования и кору.
ОБОНЯТЕЛЬНЫЙ АНАЛИЗАТОР
Рецепторы обонятельной сенсорной системы- расположены в области верхних носовых ходов. Ооонятельный эпителий находится в стороне от главного дыхательного пути, он имеет толщину 100—150 мкм и содержит рецепторные клетки диаметром 5—10 мкм, расположенные между опорными клетками (рис. 232). Общее число обонятельных рецепторов у человека — около 10 млн. На поверхности каждой обонятельной клетки имеется сферическое утолщение :—обонятельная булава, из которой выступает по 6—12 тончайших (0,3 мкм) волосков длиной до 10 мкм. Обонятельные волоски погружены в жидкую среду, вырабатываемую боуменовыми железами. Считается, что наличие волосков в десятки раз увеличивает площадь контакта рецептора с молекулами пахучих^веществ. Не исключена и активная, двигательная функция волосков, увеличивающая надежность захвата молекул пахучего вещества и контакта с ними. Булава является важным цитохимическим центром обонятельной клетки; есть основание полагать, что в ней генерируется рецепторный потенциал.
Молекулы пахучего вещества вступают в контакт со слизистой оболочкой носовых ходов, взаимодействуют со специализированными белками, встроенными в мембрану рецептора. В результате следующей за этим сложной и пока еще недостаточно изученной
цепи реакций в рецепторе генерируется рецепторный потенциал, а затем и импульсное возбуждение, передающееся по волокнам обонятельного нерва в обонятельную луковицу — первичный нервный центр обонятельного анализатора. Изменение суммарной электрической активности, регистрируемое с помощью электродов, помещенных на поверхность обонятельного эпителия, назы- мво в а ют эл е кт роол ьфа кто г р а м мой (рис. 233). Это монофазная негативная волна с амплитудой до 10 мВ и длительностью в несколько секунд, возникающая даже при кратковременном воздействии пахучего вещества. Нередко на электроольфактограмме можно видеть небольшую позитивность, предшествующую основной негативной волне, а при достаточной длительности воздействия регистрируется большая негативная волна на его прекращение (off-реакция). Иногда на медленные волны элек.троольфактограммы накладываются быстрые Осцилляции, отражающие синхронные импульсные разряды значительного числа рецепторов.
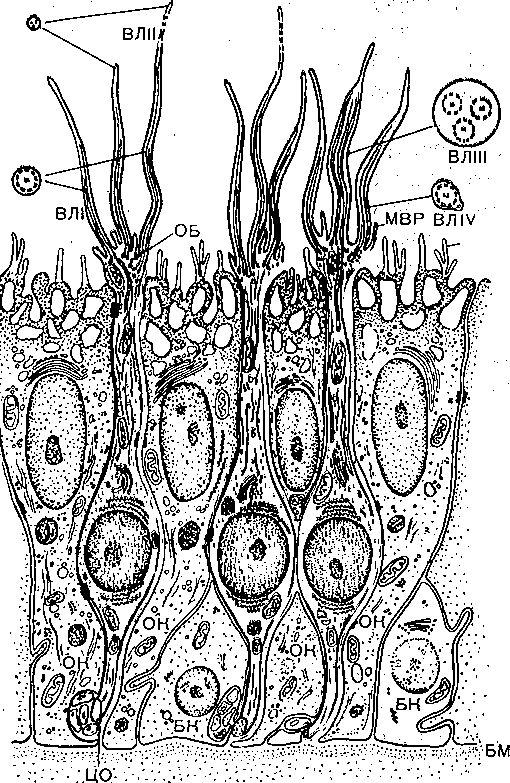
Рис. 232. Схема строения обонятельного эпителия по данным электронной микроскопии.
ОБ — обонятельная булава; ОК — опорная клетка; ЦО — центральные отростки обонятельных клеток; БК — базальная клетка; БМ — базальная мембрана; ВЛ — обонятельные волоски; МВР - микроворсипки обонятельных и МВО — микроворсинки опорных клеток.
Как показывают микроэлект родные исследования, одиночные рецепторы отвечают увеличением, частоты импульсации, которое зависит от качества и интенсивности стимула. Адаптация в обонятельном анализаторе происходит сравнительно медленно (десятки секунд или минуты) и зависит от скорости потока воздуха над обонятельным эпителием и концентрации пахучего вещества! Каждый обонятельный рецептор отвечает не на один, а на многие пахучие вещества, отдавая «предпочтение» Некоторым
из них. Считают,.что на этих свойствах рецепторов, различающихся по своей «настройке» на разные группы веществ, может быть основано кодирование раздражителей (запахов) и их опознание в центрах обонятельного анализатора. Действительно, при электрофизиологических исследованиях обонятельных луковиц выявлено, что параметры регистрируемого там при действии запахов электрического ответа-зависят от пахучего вещества. При разных запахах меняется и пространственная мозаика возбужденных и заторможенных участков луковицы. Служит ли это способом кодирования обонятельной информации, пока судить трудно.
Эфферентная регуляция активности обонятельной луковицы изучена еще недоста точно, хотя имеется много морфологических предпосылок, свидетельствующих о возмож ности таких влияний.
. Особенность обонятельного анализатора состоит^ в частности, в том, что его аффе рентные волокна не переключаются в таламусе и не переходят на противоположну* сторону большого мозга.
Выходящий из луковицы обонятельный тракт состоит из нескольких пучков, которы направляются в разные отделы переднего мозга: переднее обонятельное ядро, обоня тельный бугорок, препириформ'ную кору, периамигдалярную кору и часть ядер миндале видного комплекса. Связь обонятельной луковицы с гиппокампом, пириформной коро; и другими отделами обонятельного мозга осуществляется через несколько переключе ний. Электрофизиологические исследования и опыты на животных с условными рефлек сами показали, что наличие.значительного числа, центров обонятельного'мозга (rhinen cephalon). не является необходимым для опознания запахов. Поэтому большинств! областей, проекции обонятельного тракта можно рассматривать 'как ассоциативны центры, обеспечивающие связь обонятельной системы с другими сенсорными системам! и организацию на этой основе ряда сложных форм поведения — пищевой, оборонитель ной, половой и т. д.
Чувствительность обонятельного анализатора человека чрезвычайно велика: одш обонятельный рецептор может быть возбужден одной молекулой пахучего вещества а возбуждение небольшого числа рецепторов'приводит к возникновению ощущения В то же время изменение интенсивности действия вещества (порог различения) оцени вается людьми довольно грубо (наименьшее воспринимаемое различие в силе запах; составляет 30—60 % от его исходной концентрации). У собак эти показатели в 3—6 ра: меньше.
ВКУСОВОЙ АНАЛИЗАТОР
Вкус, так же как и обоняние, основан на хеморецепции. Вкусовые рецепторы несу информацию о. характере и концентрации веществ, поступающих в рот. Их возбуждение запускает сложную цепь реакций разных отделов мозга, приводящих к различной работе органов пищеварения или удалению вредных для организма веществ, попавши) в рот с пищей.
Рецепторы вкуса — вкусовые почки — расположены на языке, задней стенке глотки, мягком небе, миндалинах и надгортаннике. Больше всего их на кончике языка его краях и задней части. Каждая из примерно 10 ООО вкусовых почек человека состоии из нескольких (2—6) рецепторных клеток и, кроме того, из опорных клеток. Вкусовая почка имеет колбовидную форму, длина и ширина ее у человека около 7-105 м (70 мкм) она не достигает поверхности слизистой оболочки языка и соединена с полостью ртг через вкусовую пору.
в

Рис. 233. Электроольфакто грамма (ЭОГ).
а — позитивный потенциал; б - ЭОГ; в — выключение раздражите ля.
Вкусовые клетки — наиболее короткоживущие эпителиальные клетки' организма;
среднем через каждые 250 ч каждая клетка сменяется -молодой, движущейся к центру ;усовой почки от ее. периферии. Каждая из рецепторных вкусовых клеток длиной 1—20 мкм (1—2» Ю-5 м) , шириной 3—4 мкм (3—4- 10^6м) имеет на конце, обращенном просвет поры, 30—40 тончайших микроворсинок—0,1—0,2 мкм (2-Ю-7 м) длиной —2 мкм (1—2-10