Клиническая химия в диагностике и лечении. Обмен натрия и воды обмен калия лечение диуретиками гомеостаз ионов водорода
 Скачать 3.99 Mb. Скачать 3.99 Mb.
|

|
Почки Роль карбонатдегидратазы также исключительно важна в механизмах гомеостаза ионов водорода, которыми располагают почки. Ионы водорода секретируются клетками почечных канальцев в их просвет, где буферное действие обеспечивается компонентами клубочкового ультрафильтрата. В противоположность гемоглобину эритроцитов запас этих буферных систем непрерывно пополняется за счет фильтрации в почечных клубочках. По этой причине, а также учитывая тот факт, что избыток Н+ может быть удален из организма только с мочой, почки имеют наибольшее значение в процессах компенсации при хроническом ацидозе. Буферная емкость гемоглобина была бы быстро исчерпана, если бы почки не участвовали в этих компенсаторных процессах. В почках функционируют два механизма регуляции [НСОз-] в ВКЖ: реабсорбция бикарбонатов и их образование. Реабсорбция бикарбонатов — наиболее важный механизм стабилизации динамического равновесия. Двуокись углерода, необходимая для функционирования механизма карбонатдегидратазы в клетках почечных канальцев, образуется в просветах почечных канальцев из бикарбонтов ультрафильтрата. При этом общей потери ионов водорода в конечном счете не происходит. 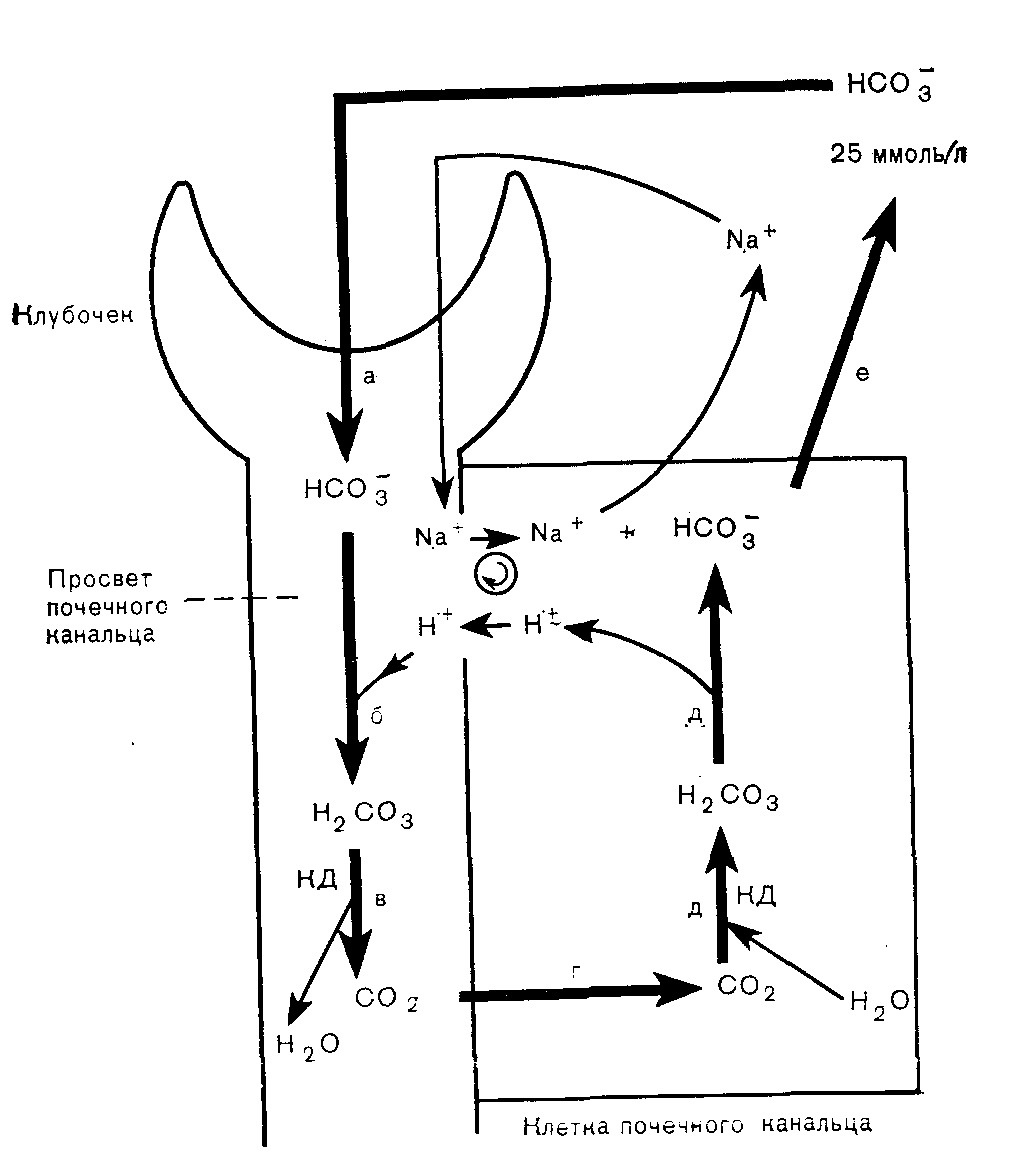 Рис 11 Реабсорбция бикарбонатов из первичной мочи в почечных канальцах. Образование бикарбонатов представляет собой очень важный для устранения ацидоза механизм, функционирующий в условиях соответствия уровня С02, стимулирующего карбонатдегидратазную реакцию в клетках почечных канальцев и в BKЖ. При этом происходят общие потери ионов водорода. Уменьшение внеклеточной [НСОз-] также стимулирует функционирование этого механизма. Реабсорбция бикарбонатов (рис. 11). В норме моча почти не содержит бикарбонатов. Поверхности клеток почечных канальцев, обращенные в их просвет непроницаемы для бикарбонатов, вследствие чего их непосредственная реабсорбция невозможна. В просвете канальцев из бикарбонатов сначала образуется С02, а в клетках канальцев эквивалентное количество С02 превращается в бикарбонаты. Функционирование этого механизма зависит от действия карбонатдегидратазы в клетках канальцев и секреции Н+ из клеток в просвет канальцев в обмен на натрий, попадающий в фильтрат вместе с бикарбонатами. На рис. 11 представлена последовательность этапов этого процесса: а) бикарбонаты проникают через клубочки в ультрафильтрат. Их концентрация в нем, как и в плазме крови, примерно 25 ммоль/л; 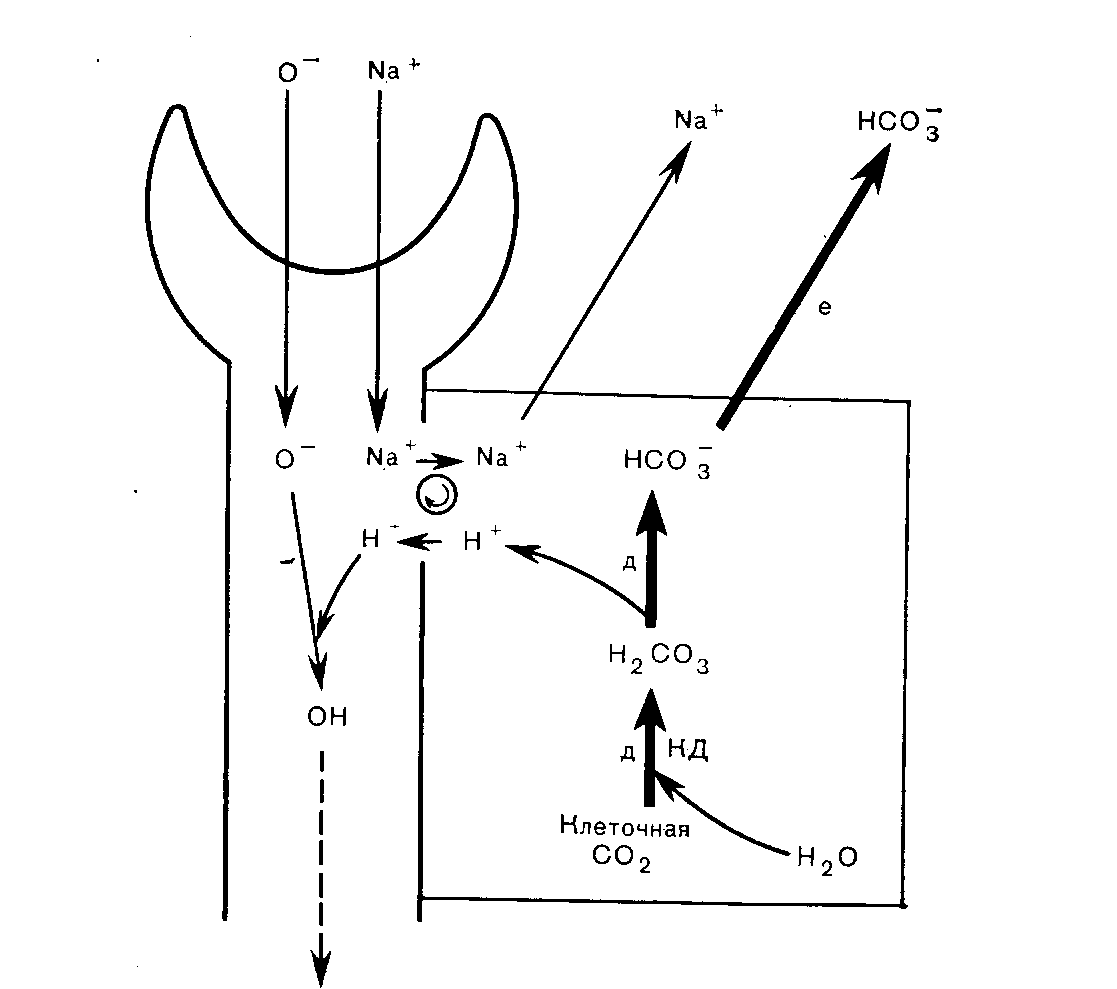 Рис. 12. Образование бикарбонатов в клетках почечных канальпев (О — основание, не являющееся бикарбонатом). б) профильтрованный бикарбонат взаимодействует с ионами водорода, секретируемыми клетками канальпев, с образованием Н2СОз; в) при диссоциации Н2СОз образуются СО2 и вода. В проксимальном отделе канальцев эту реакцию катализирует карбонатдегидратаза, локализованная на обращенных в просвет канальцев биомембранах их клеток. В дистальвых отделах нефрона, где величина рН обычно ниже, НСОз, по-видимому, диссоциирует спонтанно; г) при повышении Рсо2 в просвете канальцев происходит диффузия СОз в клетки почечных канальцев в соответствии с концентрационным градиентом; д) в условиях повышения внутриклеточной [CO2] карбонатдегидратаза катализирует ее взаимодействие с водой, приводящее вновь к образованию угольной кислоты, которая диссоциирует на Н+ и НСОз-; е) по мере секреции Н+ [инициирующей таким образом вышеописанную последовательность, начиная с пункта б] внутриклеточная концентрация НСОз- нарастает, и бикарбонат диффундирует в ВКЖ, что сопровождается реабсорбцией натрия в обмен на Д+. Благодаря этому бесконечно повторяющемуся циклу в организме сохраняются буферные системы, которые в противном случае могли бы быть выведены из него клубочковой фильтрацией. Секретируемые Н имеют своим источником внутриклеточную воду и вновь включаются в состав воды в просвете канальцев. Поскольку баланс ионов водорода не изменяется и общее количество бикарбонатов не увеличивается, этот механизм не корректирует ацидоз, но может поддерживать устойчивое состояние. Образование бикарбонатов (рис. 12). Механизм образования бикарбонатов в клетках почечных канальцев идентичен таковому реабсорбции бикарбонатов, но при его функционировании из организма выводятся Н+ и увеличивается количество НСОз-, что делает данный механизм идеальным для устранения ацидоза. Повышение Рсоз или уменьшение [НСОз-] в клетках почечных канальцев может стимулировать активность карбонатдегидратазы. При этом увеличение [CO2] является косвенным следствием повышения внеклеточного Рсо2 В противоположность эритроцитам клетки почечных канальцев в результате метаболических аэробных реакций непрерывно продуцируют CO2, которая диффундирует из клеток в ВКЖ в соответствии с концентрационным градиентом. Повышение внеклеточного Рсоз уменьшая этот градиент, замедляет указанный процесс диффузии, что приводит к повышению внутриклеточного Рсо2. Наоборот, уменьшение внеклеточной [НСОз-], увеличивая концентрационный градиент, усиливает выведение этого аниона из клеток. В норме весь профильтрованный бикарбонат реабсорбируется при участии описанного выше механизма. Как только завершается реабсорбция бикарбонатов, секреция Н+ и образование НСОз- аависят от наличия в ультрафильтрате других буферных систем (0- на рис. 12). В противоположность бикарбонатной в этих буферных системах нет соединений, способных диффундировать в клетки почечных канальцев, а ионы водорода не включаются в состав воды. Ионы водорода выводятся из организма с мочой в виде НО. В клетках бикарбонаты образуются из клеточной С02, но не бикарбонатов первичной мочи, и, следовательно, происходит общее увеличение количества бикарбонатов. Как обычно, когда с мочой экскретируется 1 ммоль Н+, в ВКЖ вместе с натрием поступает 1 ммоль НСОз-. Этот механизм очень похож на функционирующий в эритроцитах, но в отличие от последних клетки почечных канальцев не перемещаются между легкими и периферическими тканями. На них поэтому воздействует относительно постоянное Рсо2 Связанное с секрецией ионов водорода образование бикарбонатов приобретает очень важное значение при ацидозе, когда оно стимулируется уменьшением внеклеточной [НСОз-] (метаболический ацидоз) или повышением внеклеточного Рсо2 (дыхательный ацидоз). Буферные системы мочи. В образование бикарбонатов с секрецией Н+, помимо бикарбонатной, вовлечены и другие буферные системы. Среди них наиболее важны фосфатная и аммонийная. Фосфатная буферная пара. При рН 7,4 фосфаты клубочкового ультрафильтрата преимущественно представлеяы ионами двузамещенных фосфатов (HP04=), которые могут, присоединяя Н+, становиться ионами однозамещенных фосфатов (Н2Р04-). Величина рК пары равна 6,8. Даже при небольшом ацидозе из солей костной ткани высвобождается больше кальция и фосфатных ионов, чем при нормальных значениях рН. При этом потребности в увеличении экскреции Н+ с мочой соответствует повышение буферной емкости клубочкового фильтрата, что обусловлено увеличением содержания в нем фосфатов, высвобождаемых из костной ткани. Роль аммиака. По мере увеличения секреции Н+ все большее количество двузамещенных фосфатов превращается в однозамещенные до тех пор, пока (при рН менее 5,5) основная масса фосфатов не будет в этой форме. При тяжелом ацидозе важное значение приобретает образование бикарбонатов, но фосфаты мочи не могут поддерживать постоянную непрерывную секрецию ионов водорода. Преобладающий количественно в моче анион хлорид не может оказать буферное действие на ионы водорода (хлористоводородная или соляная кислота ионизирована почти полностью). По мере того как моча становится более кислой, в ней нарастает содержание ионов аммония. Повидиому, амиак мочи способствует дальнейшей секреции ионов водорода и образованию бикарбонатов. В клетках почечных канальцев имеется фермент глутаминаза, катализирующий гидролиз конечной аминогруппы глутамина (GluCONH2) с образованием глутамина (GluCOO-) и иона аммония, 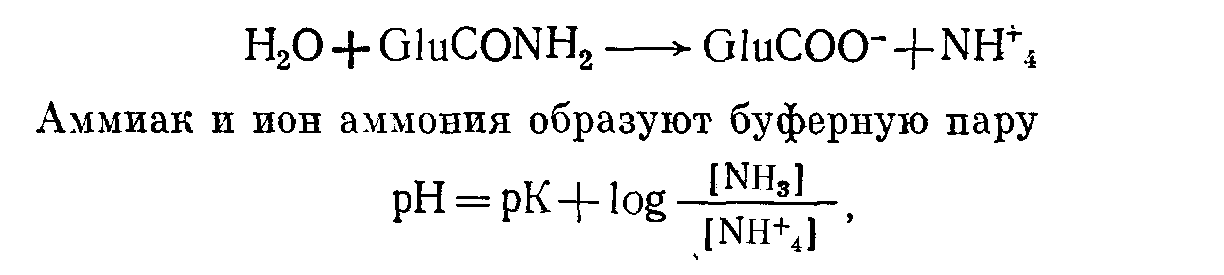 но значение рН для этой системы равно примерно 9,8, а при рН 7,4 равновесие резко смещено к NH4+. Диффузия NH3 из клеток в мочу происходит со значительно большей скоростью, чем NH4+. При кислой реакции мочи обратная диффузия NH3 не происходит, так как он жадно взаимодействует с секретируемыми (прп участии карбонатдегидратазы) ионами Н+. В этой ситуации возможна экскреция ионов водорода в виде хлорида аммония. Поэтому при сильном ацидозе образование бикарбонатов может продолжаться даже после того, как буферная емкость фосфатов истощена. Диссоциацию NH4+ в клетках обеспечивает выведение NH3 с мочой. Однако эта диссоциация высвобождает Н+ в клетках. Казалось бы, нет никакого смысла в буферном действии в отношении сек рйтируемых Н+ в моче, если одновременно Н+ образуются в клетках. Объяснение этих явлений возможно на основе изучения превращений глутамата (GluCOO-), образующегося одновременно с ионам аммония. Глутамат после дезаминирования до аоксоглутарата может превращаться в глюкозу путем глюконеогенеза — процесса, при котором используется эквивалентное количество Н+. Таким образом, высвобождаемые в клетках Н+ могут включаться в состав глюкозы. 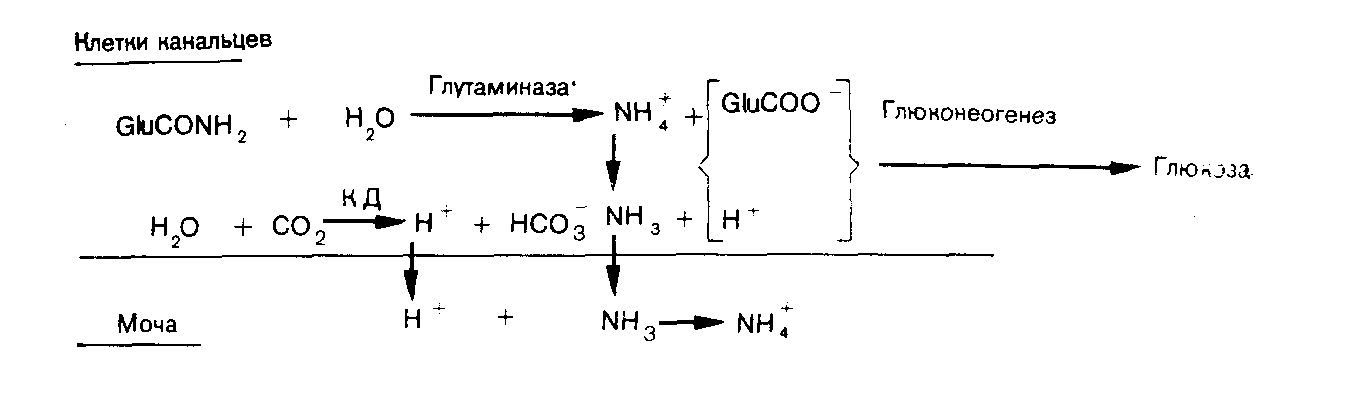 Как обычно, конечным результатом является увеличение НСОз-. При ацидозе повышается активность глутаминазы и усиливается глюконеогенез. Образование бикарбонатов в желудочнокишечном тракте В клетках слизистой оболочки кишечника бикарбонаты образуются при участии карбонатдегидратазы. Они могут поступать либо в ВКЖ, либо в просвет кишечника, но в любом случае необходимым условием является перекачивание Н+ в противоположном направлении. Электрохимическая нейтральность обеспечивается либо обменом Na+ на Н+ при участии механизма, обратного функционирующему в почках, либо перемещением Сl- вместе с Н+. Секреция кислоты в желудке. Париетальные клетки желудка секретируют Н+ вместе с Сl- в его полость. Как только Н+ Сl- поступают в полость желудка, увеличивается [НСОз-] в ВКЖ, что объясняет явление щелочного прилива после приема пищи. У здорового человека это явление быстро корректируется секрецией бикарбонатов по мере того, как пищевые массы перемещаются по желудочнокишечному тракту. Этот механизм важен для объяснения развития метаболического алкалоза, обусловленного стенозом пилорической части желудка. Секреция бикарбоната натрия клетками поджелудочной железы и желчных протоков. Содержимое двенадцатиперстной кишки является щелочным, поскольку поступающий в него через общий желчный проток секрет содержит много бикарбонатов. Секреция бикарбоната натрия клетками поджелудочной железы и желчных 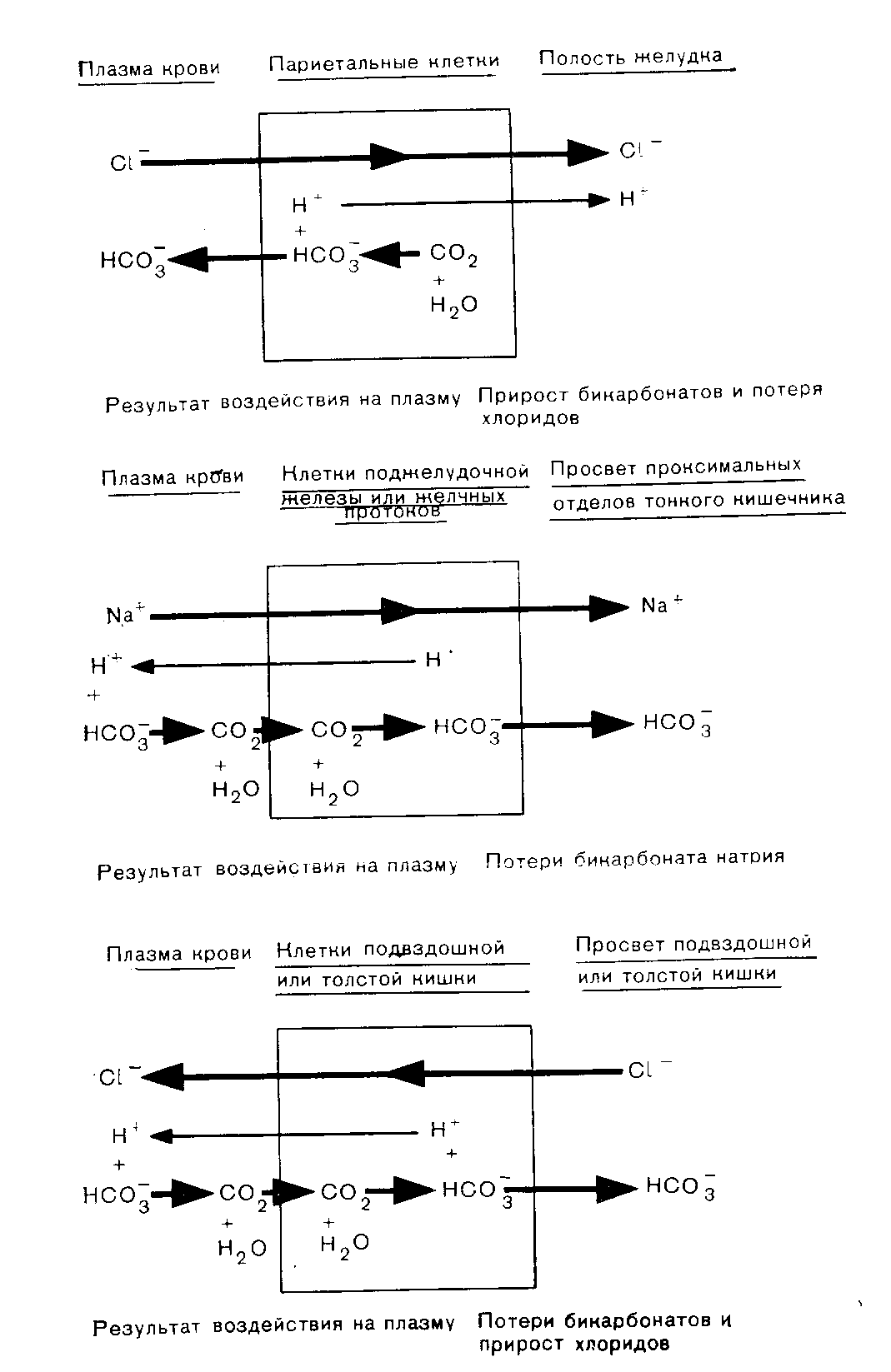 Рис. 13. Возможные механизмы превращений бикарбонатов в желудочнокишечном тракте. протоков представляет собой процесс, обратный его реабсорбции в клетках почечных канальцев. Функционирующие в поджелудочной железе и желчных протоках механизмы стимулируются местным повышением Pco2 когда Н+ перекачиваются из железистых клеток в ВКЖ и реагируют с НСОз-, образуемым париетальными клетками желудка. Эти процессы аналогичнь! стимуляции образования бикарбонатов в почках при повышении Рсоа в просвете канальцев. Потеря организмом большого количества дуоденального сока может вызвать истощение резерва бикарбонатов. Секреция бикарбонатов и реабсорбция хлоридов клетками желуДочнокишечного тракта. По мере продвижения содержимого жеудочнокишечного тракта по кишечнику в его просвет поступают бикарбонаты, тогда как хлориды перемещаются в противоположном направлении под действием механизма, обратного функционирующему в слизистой оболочке желудка. Таким образом корректируются происходящие в желудке потери хлоридов в увеличение содержания бикарбонатов. Предпочтительная реабсорбция хлоридов мочи при функционировании этого механизма объясняет факт развития гиперхлоремического ацидоза после трансплантации мочеточника в подвздошную кишку или в толстый кишечник. На рис. 13 представлены в общем виде механизмы превращений бикарбонатов в желудочнокишечном тракте. Итак, СО2 играет ведущую роль в гомеостазе ионов водорода. Несмотря на казалось бы неблагоприятную для функционирования в физиологических условиях величину рК бикарбонатной буферной системы, тенденция Н2СОз к образованию газообразной С02, парциальное давление которой благодаря регулирующим воздействиям дыхательного центра и легких поддерживается в диапазоне 5,3 кПа, а также способность клеток почечных канальцев и эритроцитов поддерживать [НСОз-], равной 25 ммоль/л при данном Рсоа» обеспечивают стабилизацию рН на уровне, превосходящем величину рК этой системы. Нарушения функций почек или легких приводят к расстройствам регуляции внеклеточного рН. Ингибирование активности карбонатдегидратазы (с. 124) снижает образование бикарбонатов в эритроцитах и клетках почечных канальцев, угнетает реабсорбцию бикарбонатов из клубочкового фильтрата и приводит к истощению резерва бикарбонатов. Помимо бикарбонатов и гемоглобина, к числу буферных систем организма относятся белки плазмы крови и тканей. Внутриклеточные белки имеют важное значение для местных буферных аффектов, но роль белков плазмы крови в гомеостазе ионов водорода весьма мала. Фосфатные буферные системы, имеющие рК 6,8, очень важны для обеспечения буферных эффектов в моче, где их концентрация может достигать 25 ммоль/л. В плазме крови концентрация фосфатов составляет лишь около 1 ммоль/л, и они не играют важной роли в буферном действии крови. НАРУШЕНИЯ ГОМЕОСТАЗА ИОНОВ ВОДОРОДА В нарушениях гомеостаза ионов водорода участвуют бикарбонатные буферные пары. При дыхательных нарушениях первичными являются изменения содержания СО2 но при так называемых метаболических расстройствах прежде всего изменяется [НСОз-], тогда как отклонения от нормы в содержании CО2 являются вторичными. Измерения, применяемые для оценки баланса ионов водорода Измерения рН крови указывают лишь на наличие явного ацидоза или алкалоза. Если имеются отклонения от нормы величины рН крови, то причиной этого может быть нарушение механизма регуляции содержания CО2 (включающего легкие и дыхательный центр) или дисбаланс между использованием бикарбонатов для обеспечения буферных эффектов и их реабсорбцией и образованием в клетках почечных канальцев и в эритроцитах. Однако нормальная величина рН крови не исключает возможности нарушения перечисленных процессов; она может поддерживаться компенсаторными механизмами. Для оценки этих факторов необходимо определение содержания компонентов бикарбонатной буферной системы. Концентрацию растворенной СО2 рассчитывают, умножая измеренное Рсо2 на константу растворимости данного газа (0,23, если Рсо2 выражено в кПа; 0,03, если оно выражено в миллиметрах ртутного столба). рН и Рсо2. Для измерения этих величин следует использовать артериальную кровь, так как имеется значительная артериовенозная разница в величинах рН и Рсо2. Поскольку на результаты определений Рсог некоторыми методами может влиять присутствие эритроцитов, для исследования требуется цельная (гепаринизированная) кровь. Важное значение имеет методика взятия крови, подробно описанная на с. 135. Измерение [НСОз-] в крови. Для определения концентрации бикарбонатов в циркулирующей крови можно применить один из следующих двух методов. 1. Общая концентрация бикарбонатов (Тсо2) в плазме крови. Это, по-видимому, наиболее часто и легко измеряемый показатель гомеостаза ионов водорода. Для обеспечения надежных результатов требуется получить достаточное количество плазмы крови (с. 133). Если нет необходимости определять рН и Рсо2, можно использовать для анализов венозную кровь, что является определенным преимуществом данного метода, позволяющего оценить суммарное содержание в плазме крови бикарбонатов, угольной кислоты и растворенной С02. При рН 7,4 отношение [НСОз-] к концентрациям двух других указанных компонентов составляет примерно 20:1, а при рН 7,1—10:1. Таким образом, если Tcog равна 21 ммоль/л, то доля [НСОз-] при рН 7,4 составляет 20 ммоль/л и при рН 7,1 — несколько более 19 ммоль/л. На долю Н2СОз+СО2 приходится соответственно лишь 1 моль/л и несколько меньше 2 ммоль/л. Следовательно, Тсо2 дает адекватное представление о концентрации бикарбонатов в плазме крови. 2. Фактическая концентрация бикарбонатов. Для этого анализа берут цельную артериальную кровь. Рассчитывают по уравнению Гендерсона—Гассельбалха, используя измеренные величины 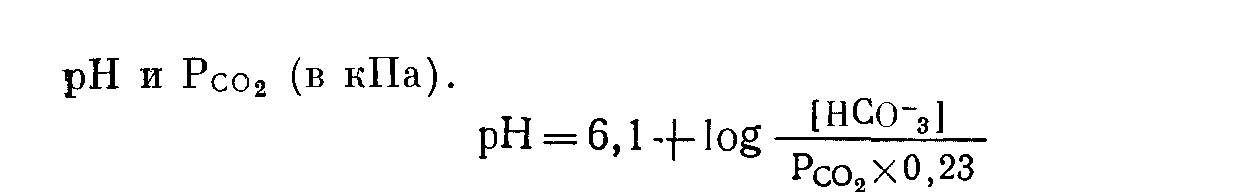 Определяемая таким путем величина характеризует [НСОз-] цельной крови и по причинам, рассмотренным выше, обычно хорошо коррелирует с Тсо2 плазмы крови. Если известны два других параметра, то целесообразнее использовать именно этот метод. Ацидоз Ацидоз обусловлен уменьшением отношения [НСОз-]: Рсо2 в ВКЖ. При метаболическом (нереспираторном) ацидозе первичным нарушением в бикарбонатной буферной системе является уменьшение [НСОз-]. При респираторном ацидозе первичным нарушением в бикарбонатной буферной системе является повышением Рсо2. Как при метаболическом, так и при респираторном ацидозах отношение [НСОз-]: Рсог и, следовательно, рН можно нормализовать, изменяя концентрацию другого компонента буферной пары в направлении, соответствующем таковому первичного нарушения. Эта компенсация может быть частичной или полной. При метаболическом ацидозе компенсаторный сдвиг заключается в снижении Рсо2; при респираторном ацидозе — в увеличении [НСОз-]. При полностью компенсированном ацидозе величина рН в пределах нормы. Однако содержание других компонентов, фигурирующих в уравнении Гендерсона—Гассельбалха, отклоняется от нормы. Полная нормализация всех этих параметров возможна только после устранения первичного нарушения. |