Программа блока гетерофункциональные углеводороды
 Скачать 6.9 Mb. Скачать 6.9 Mb.
|

|
Отдельные представители трисахаридов Принципы построения трисахаридов такие же, как и дисахаридов, т. е. трисахариды могут быть как восстанавливающие, так и невосстанавливающие. Многие из трисахаридов содержатся в растениях. Из трисахаридов наиболее изучены рафиноза, генцианоза и целлотриоза. Рафиноза (мелитриоза) С18Н32О16. Встречается во многих растениях, в частности в семенах хлопчатника и в сахарной свекле; не имеет сладкого вкуса. Интересно, что при хранении свеклы содержание рафинозы в ней возрастает. 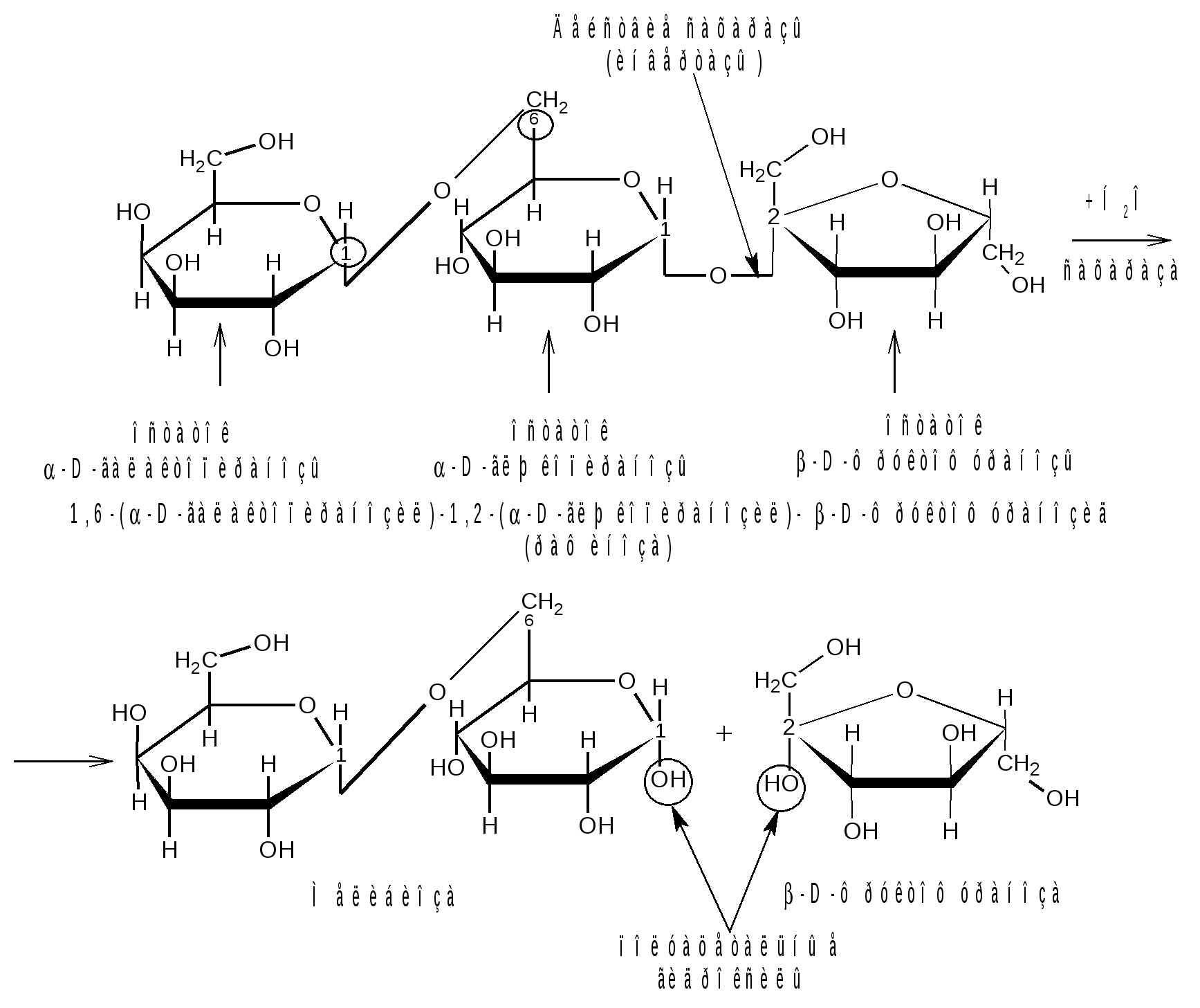 При действии фермента -галактозидазы, содержащегося в эмульсине, рафиноза расщепляется на галактозу и сахарозу: 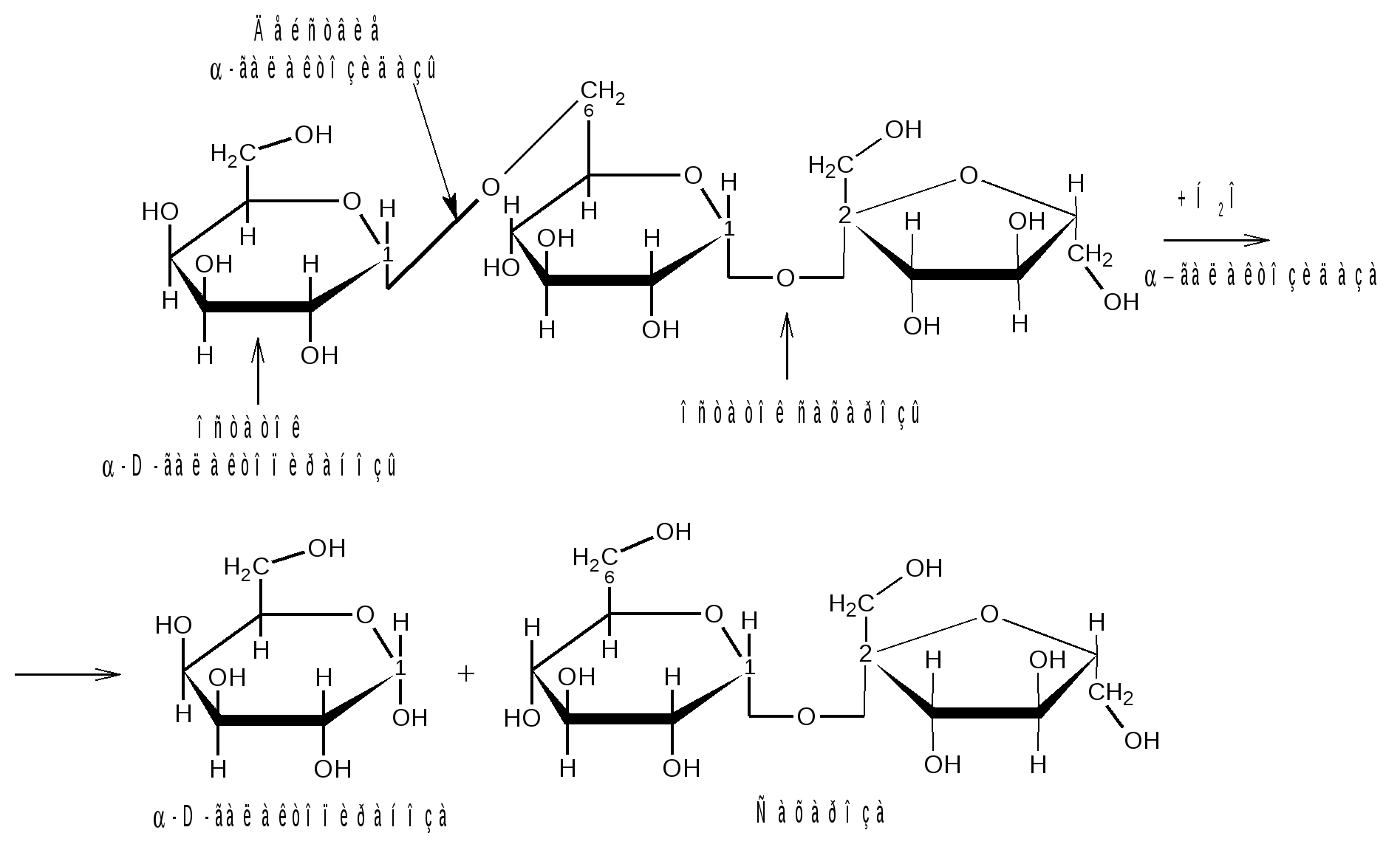 Генцианоза С18Н32О16. Содержится в корнях горечавки, относится к невосстанавливающим трисахаридам. При полном гидролизе образует две молекулы D-глюкозы и одну молекулу D-фруктозы (кислотный гидролиз). Ферментативный гидролиз генцианозы идет по двум направлениям. Под действием фермента инвертазы (сахаразы) или разбавленной серной кислоты она расщепляется на -D-фруктофуранозу и дисахарид генциобиозу, а под действием фермента -глюкозидазы образуются -D-глюкопираноза и сахароза: 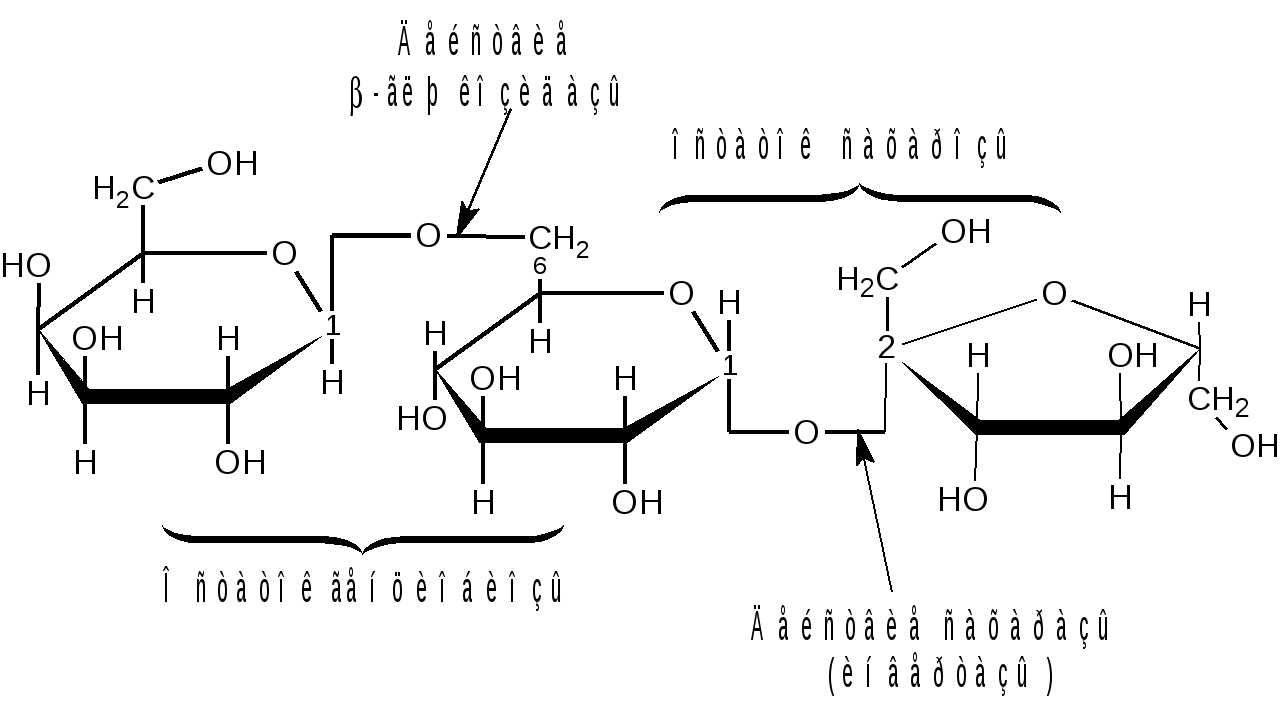 Таким образом, генцианоза представляет собой 1,6-(-D-глюко-пиранозил)-1,2-(-D-глюкопиранозил)--D-фруктофуранозид. Целлотриоза С18Н32О16 состоит из трех молекул -D-глюкопиранозы по типу целлобиозы и является, следовательно, 1,4-(-D-глюкопиранозил)-1,4-(-D-глюкопиранозил)--D-глюкопиранозой. Целлотриоза относится к восстанавливающим олигосахаридам, в ее молекуле имеется один полуацетальный гидроксил и поэтому она восстанавливает фелингову жидкость, водные растворы целлотриозы мутаротируют. Молекулы глюкозы в составе целлотриозы соединены между собой с помощью двух -1,4-гликозидных связей. Целлотриоза расщепляется ферментом -глюко-зидазой с образованием трех молекул глюкозы: 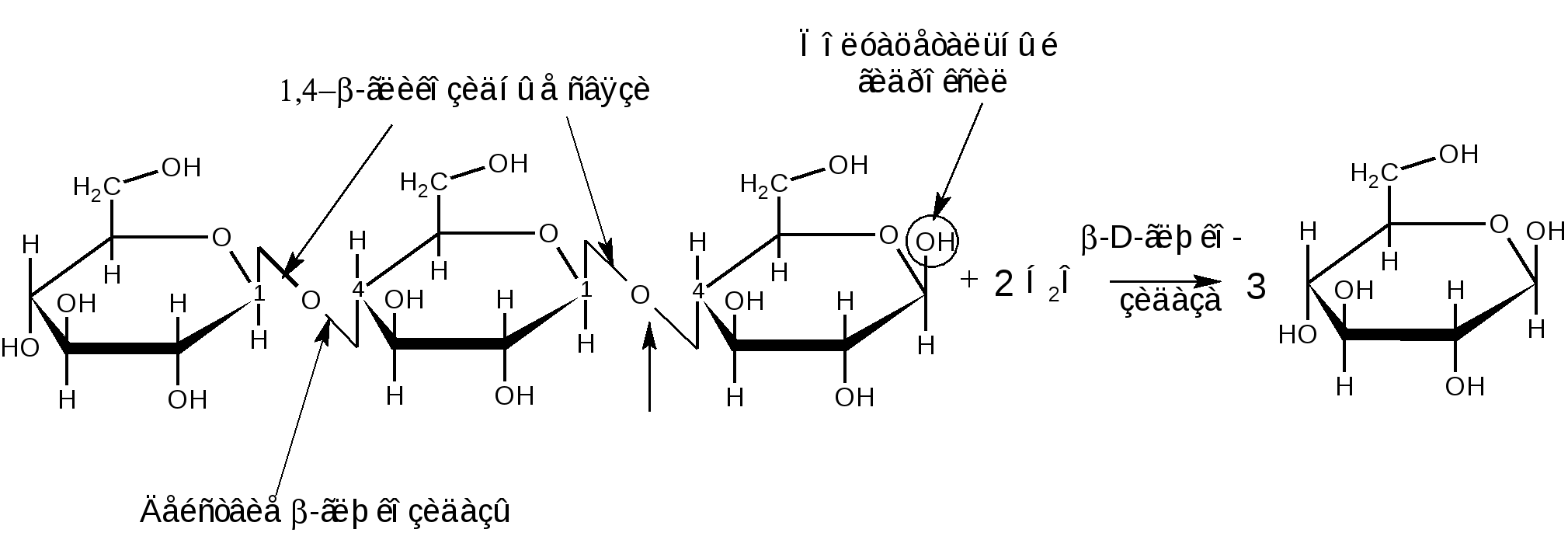 Природные гликозиды Гликозиды – это простые или сложные эфиры моносахаридов и олигосахаридов, образованные с участием полуацетального гидроксила. Гликозиды чрезвычайно распространены в животном и особенно в растительном мире. При гидролизе природных гликозидов получаются моносахариды и олигосахариды, а также неуглеводный компонент (агликон). Агликоны представляют собой гидроксисоединения жирного и ароматического рядов. Природные гликозиды наиболее часто классифицируют по характеру их агликонов. Так, известны фенолгликозиды, содержащие в качестве агликонов фенолы и фенолоспирты; антрахинонгликозиды, в которых агликоном служат производные антрахинона; флавонгликозиды, дающие при гидролизе призводные флавона и т. д. Особую группу гликозидов образуют дубильные вещества. Многие гликозиды применяются в медицине. Огромное значение имеют нуклеозиды, являющиеся N-гликозидами и имеющие в качестве агликонов пуриновые и пиримидиновые основания Нуклеозиды являются структурными компонентами биологически важных веществ: нуклеотидов, нуклеиновых кислот и нуклеопротеидов. Подавляющее большинство известных природных гликозидов являются -гликозидами и поэтому способны подвергаться гидролитическому расщеплению под влиянием -гликозидаз семян миндаля. Гликозид амигдалин содержится в семенах горького миндаля, в косточках персика, абрикосов и слив, вишен, яблок, груш, в листьях лавровишни и т. д. Под действием кислот амигдалин распадается на две молекулы глюкозы, одну молекулу бензойного альдегида и одну молекулу синильной кислоты: 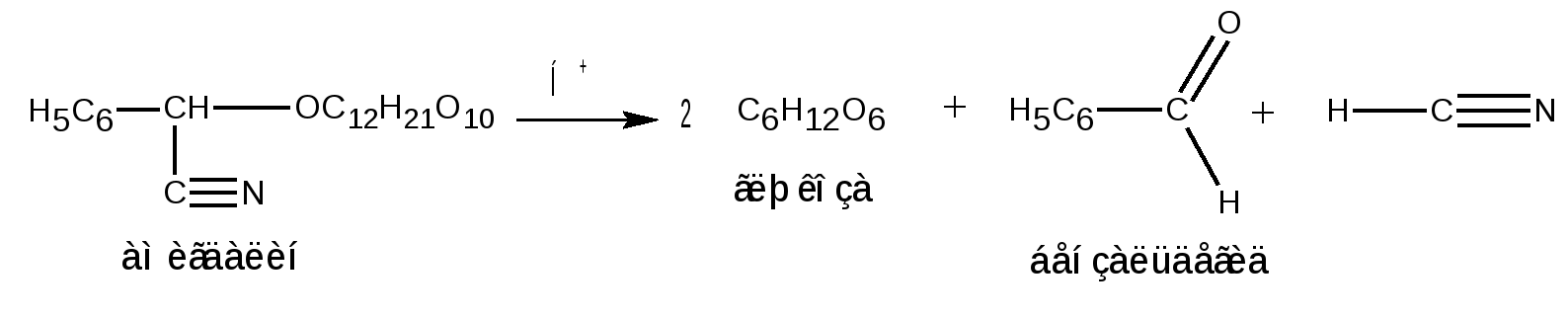 Глюкоза находится в амигдалине в виде дисахарида генциобиозы. Высокоактивная -гликозидаза, которая содержится в эмульсине – ферментном препарате из миндаля, довольно быстро расщепляет обе -гликозидные связи амигдалина, но одна из них – соединяющая гликозидные остатки в генциобиозе, расщепляется быстрее: 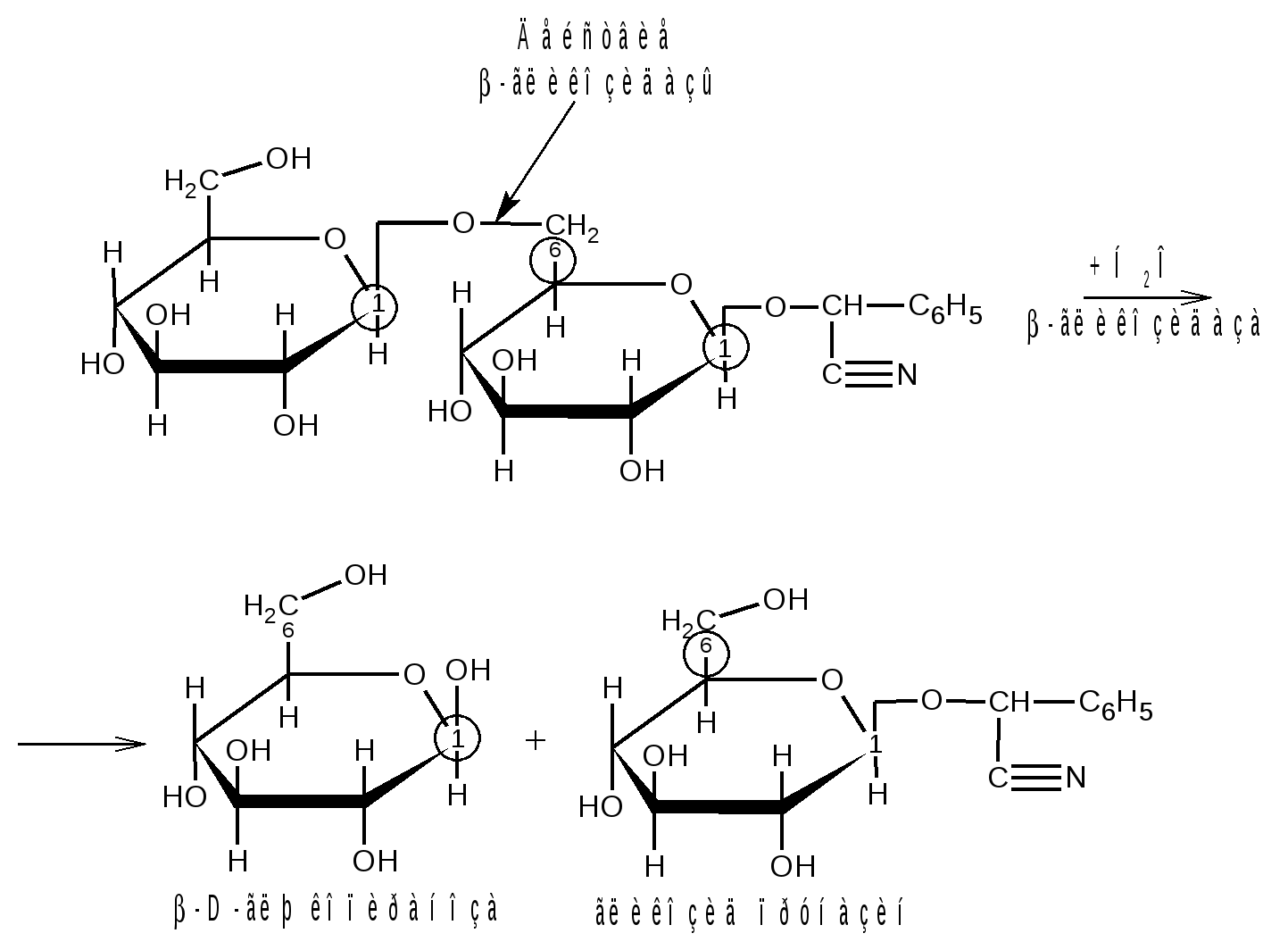 Синигрин – гликозид семян горчицы и корней хрена. Калиевая соль синигрина под действием фермента, называемого мирозином, распадается на глюкозу, аллилгорчичное масло и гидросульфит калия: 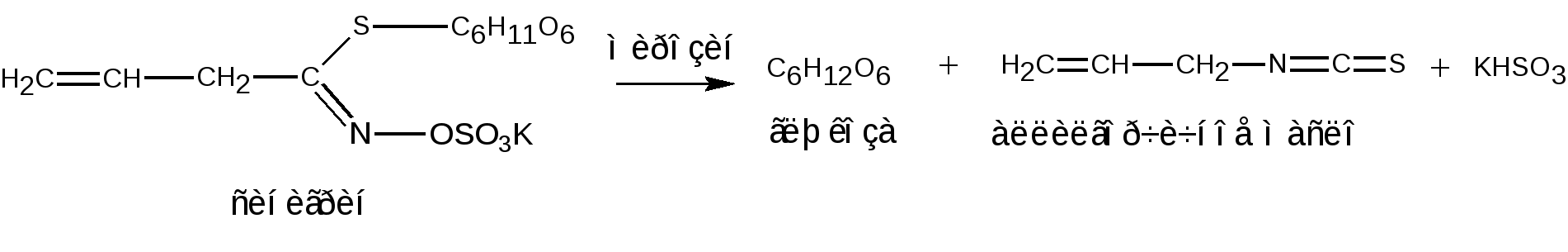 В этом гликозиде полуацетальный кислород в молекуле глюкозы заменен атомом серы. Можно произвести расщепление синигрина таким образом, чтобы атом серы остался в молекуле глюкозы и получить 1-тио-глюкозу: 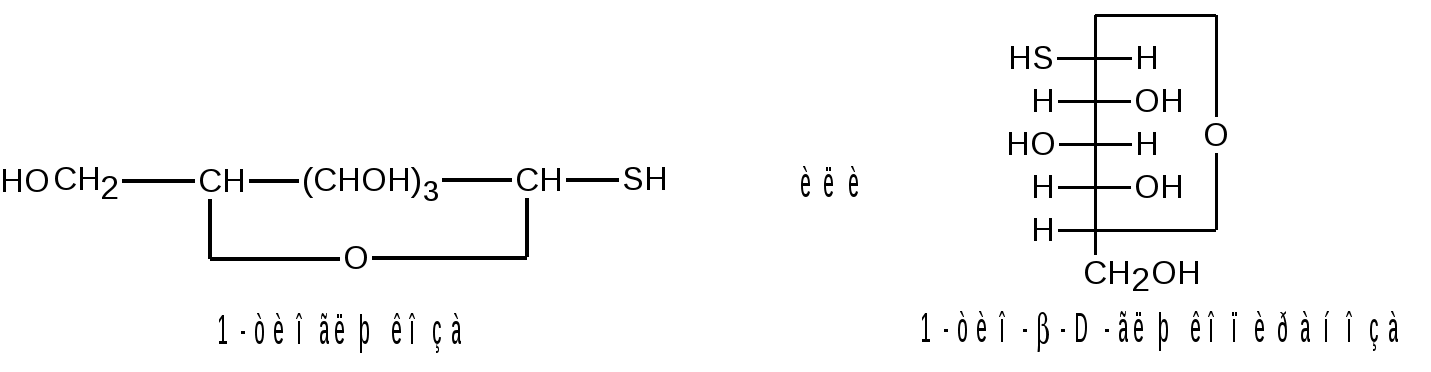 1.2.3 НЕСАХАРОПОДОБНЫЕ ПОЛИСАХАРИДЫ Строение высших полиоз Несахароподобные полиозы – высокомолекулярные соединения, содержащие сотни и тысячи остатков моносахаридов. Остатки моноз связаны между собой кислородными мостиками, образованными за счет полуацетального (гликозидного) гидроксила предыдущего остатка моносахарида со спиртовым гидроксилом последующего. Таким образом, полимерные цепи полиоз по своему строению представляют собой полигликозиды (полиацетали). В полисахаридах растительного происхождения в основном образуются (1 4)- и (1 6)-гликозидные связи, а в полисахаридах животного и бактериального происхождения дополнительно имеются также (1 3)- и (1 2)-гликозидные связи. На конце цепи находится остаток восстанавливающего моносахарида. Поскольку доля концевого остатка относительно всей макромолекулы невелика, то полиозы проявляют очень слабые восстановительные свойства. Гликозидная природа полисахаридов обусловливает их гидролиз в кислой и высокую устойчивость в щелочной средах. Полный гидролиз приводит к образованию моносахаридов или их производных, неполный – к ряду промежуточных олигосахаридов, в том числе и дисахаридов. Полисахариды имеют большую молекулярную массу. Им присущ характерный для высокомолекулярных веществ более высокий уровень структурной организации макромолекул. Наряду с первичной структурой, т. е. определенной последовательностью мономер-ных остатков, важную роль играет вторичная структура, определяемая пространственным расположением макромолекулярной цепи. Полисахаридные цепи могут быть разветвленными или неразветвленными (линейными). Для полисахаридов используется общее название – гликаны. Гликаны могут быть гексозанами или пентозанами, т.е. состоять, соответственно, из гексоз или пентоз. В зависимости от природы моносахарида различают глюканы, маннаны, галактаны, ксиланы, арабаны и т. п. Классификация высокомолекулярных полисахаридов. По строению высшие полиозы делят на следующие группы:
К гомополисахаридам относятся многие полисахариды растительного (крахмал, целлюлоза, пектиновые вещества), животного (гликоген, хитин) и бактериального (декстраны) происхождения. Гетерополисахариды изучены меньше, однако они играют важную биологическую роль. Гетерополисахариды в организме связаны с белками и образуют сложные надмолекулярные комплексы. Функции углеводов в живых организмах многообразны. Структурные полисахариды, играющие опорную роль в организмах растений и животных, – целлюлоза и пектиновые вещества растений, хитин насекомых и грибов. Резервные полисахариды, являющиеся источником энергии для живых организмов, – крахмал, гликоген, инулин, лихенин. Энергия окисления углеводов используется в различных обменных процессах; атомы углерода необходимы для биосинтеза белков, нуклеиновых кислот, липидов и др. соединений. Наличие углеводных меток на поверхности клеток позволяет живому организму отличать свои клетки и макромолекулы от чужих. Свойственна углеводам и защитная функция от разрушающего воздействия внешней среды или чужих ферментов, а также рецепторная роль – связывание гормонов, токсинов и других продуктов метаболизма. В соединениях с белками и липидами углеводы образуют сложные высокомолекулярные комплексы, представляющие основу природных биополимеров, в том числе нуклеиновых кислот, несущих наследственную информацию всех живых организмов. 1.2.3.1 ГОМОПОЛИСАХАРИДЫ Крахмал представляет собой не однородное вещество, а смесь полисахаридов, отличающихся не только степенью полимеризации, но и строением. Изучение химических превращений крахмала было начато в 1814 г. в России К.С.Кирхгофом, впервые осуществившим кислотный и ферментативный гидролиз крахмала и выделившим из гидролизата глюкозу. Крахмал является одним из важнейших продуктов фотосинтеза, образующимся в зеленых листьях растений в виде так называемых первичных зерен. Затем он расщепляется на моносахариды или их фосфорнокислые эфиры и переносится в другие части растений, например, в клубни картофеля или зерна злаков. Здесь вновь происходит отложение крахмала в виде зерен. Крахмальные зерна различных растений имеют неодинаковую величину, форму и строение, так что по внешнему виду можно определить их происхождение. Наиболее высоко содержание крахмала в зернах риса (62–82 %), кукурузы (65–72 %), пшеницы (57–75 %) и клубнях картофеля (12–24 %), из которых обычно и получают крахмал в производственной практике. Крахмал представляет собой белый аморфный порошок («картофельная мука»). Высушенный при 100–110 С крахмал очень гигроскопичен. В холодной воде он нерастворим, а в горячей набухает и некоторая часть его постепенно растворяется. Широко известна цветная реакция крахмала с йодом, при этом возникающее интенсивное синее окрашивание исчезает при кипячении раствора и вновь появляется при охлаждении его. Крахмал практически не обладает восстанавливающей способностью: он не восстанавливает ни фелингову жидкость, ни гидроксид меди, ни аммиачный раствор оксида серебра. Однако более чувствительные окисляющие реактивы (динитросалициловая кислота) дают возможность обнаружить в крахмале ничтожное количество свободных альдегидных групп – одну на сотни и тысячи глюкозных остатков. При быстром нагревании крахмала за счет содержащейся в нем влаги (10–20 %) происходит гидролитическое расщепление макромолекулярной цепи на более мелкие осколки и образуется смесь полисахаридов, называемых декстринами. Декстрины растворяются в воде лучше, чем крахмал. Такой процесс расщепления крахмала осуществляется при хлебопечении. Крахмал муки, превращенный в декстрины, легче усваивается вследствие большей растворимости. Еще быстрее декстринизация протекает в присутствии кислот или ферментов. По мере убывания величины молекулы декстрины дают с йодом окрашивание в сине-фиолетовый, фиолетовый, красно-фиолетовый, красно-оранжевый, оранжевый и желтый цвет. Как при ферментативном, так и при достаточно продолжительном кислотном расщеплении крахмала гидролиз обычно не заканчивается образованием декстринов, ферменты пищеварительных соков при температуре около 40 С гидролизуют крахмал до глюкозы; так же действуют и минеральные кислоты при длительном кипячении. Схема постепенного гидролиза крахмала кислотным или ферментативным способом в упрощенном виде выражается следующим образом: (С6Н10О5)n (С6Н10О5)m С12Н22О11 С6Н12О6. Крахмал, по строению входящих в его состав полисахаридов, можно разделить на две основные фракции: амилозу и амилопектин. Содержание этих фракций в крахмале различных растений различно: 15–25 % амилозы и 75–85 % амилопектина. Полисахариды амилозы имеют неразветвленную (или очень мало разветвленную) цепь -D-глюкопиранозных остатков, связанных -1,4-гликозидными связями, т. е. дисахаридным фрагментом амилозы является мальтоза: 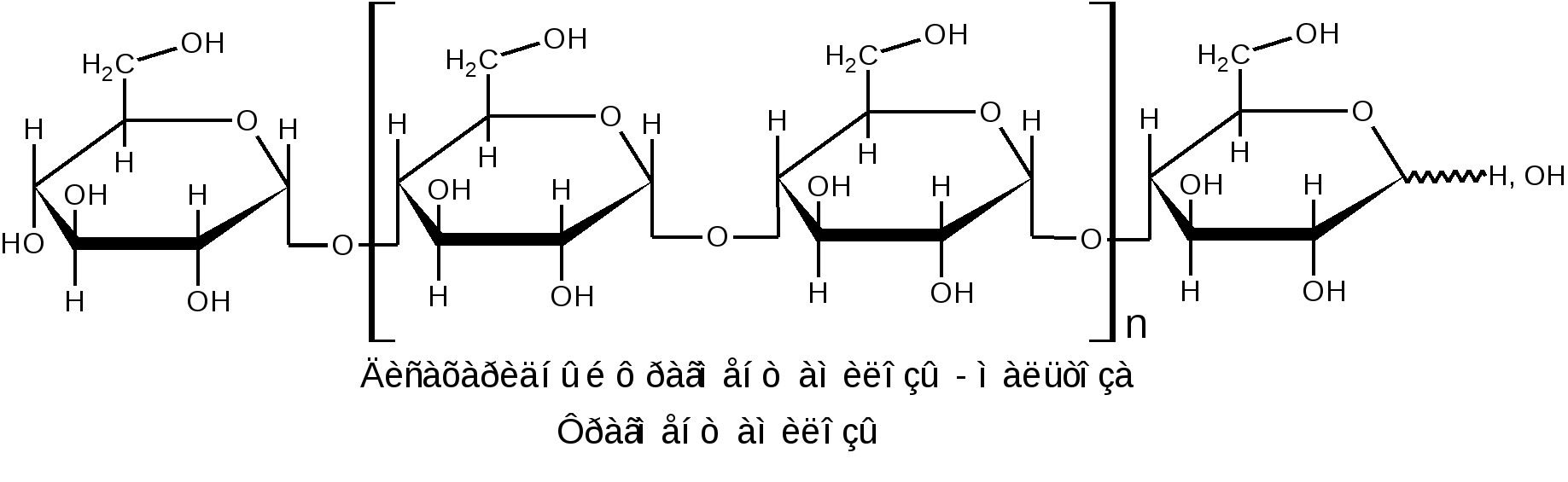 Из формулы видно, что каждое элементарное звено полимерной цепи кроме конечных звеньев содержит по три гидроксила: у второго, третьего и шестого атомов углерода. Свободный полуацетальный гидроксил находится только на конце цепи, что объясняет незначительную восстанавливающую способность полисахарида. По данным рентгеноструктурного анализа пространственная конформация цепной макромолекулы амилозы имеет форму спирали. Такая форма обусловлена конфигурацией -гликозидных связей. На каждый виток спирали приходится 6 остатков глюкопиранозы. Во внутренний канал спирали могут входить соответствующие по размеру молекулы, например, молекулы йода образуют комплексы, называемые соединениями включения. Комплекс амилозы с йодом имеет синий цвет. Это используется в аналитических целях для открытия как крахмала, так и йода (йодкрахмальная проба). Пространственное расположение атомов и звеньев в цепной макромолекуле амилозы дано ниже. 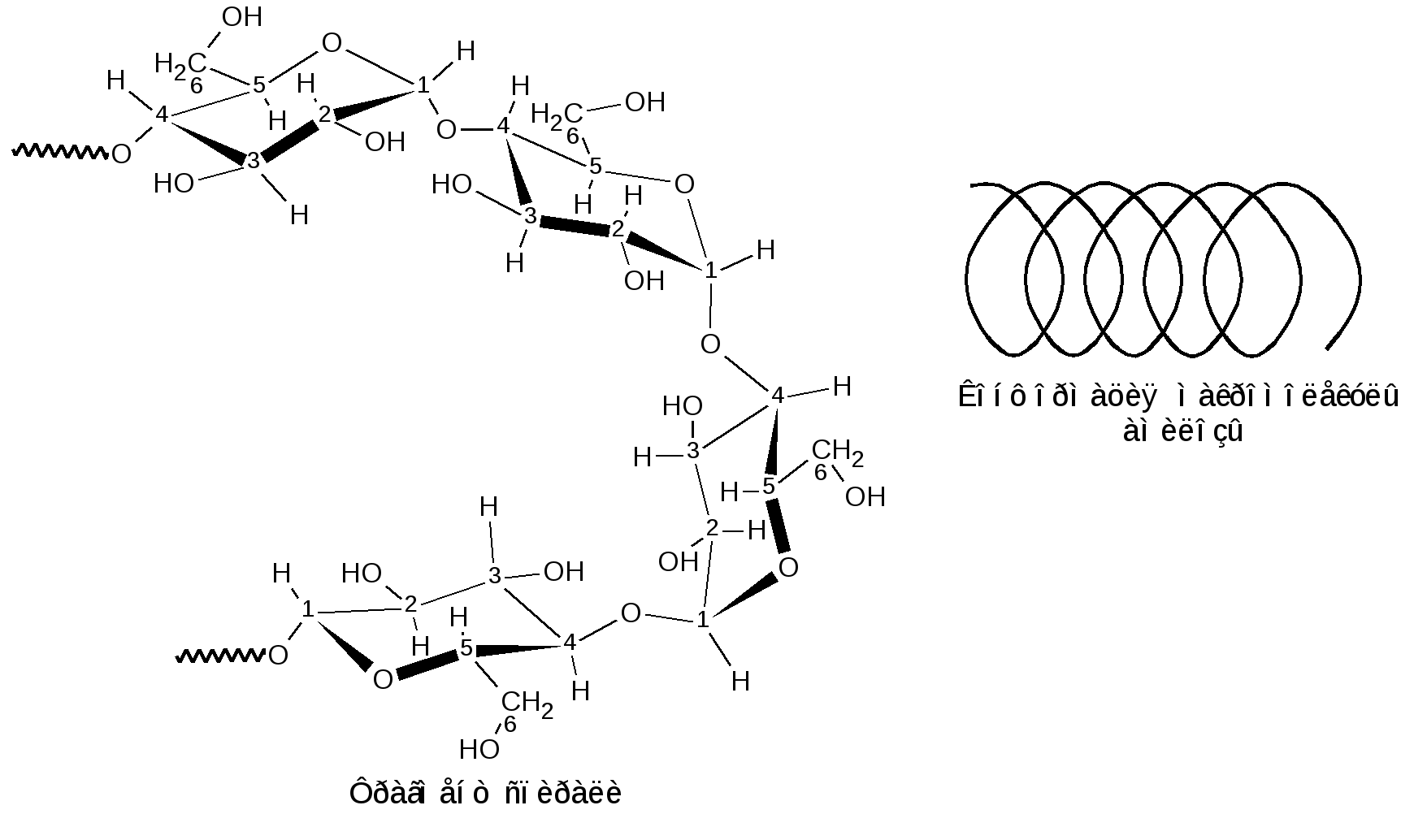 Молекулярная масса амилоз, определяемая различными методами, составляет 160 тыс. до 1 млн. (и даже выше), что соответствует наличию в моле куле от 1000 до 6000 и более глюкозных остатков. Амилоза может быть полностью расщеплена ферментом -амилазой до мальтозы. Водные растворы амилозы очень нестойки. Через некоторое время она выпадает в осадок. Это явление носит название ретроградации. Амилопектин в отличие от амилозы имеет разветвленное строение. В цепи D-глюкопиранозные остатки, как в амилозе, связаны -1,4-гликозидными связями, а в точках ответвления присутствуют -1,6-гликозидные связи. Между точками ответвлений располагаются 20–25 глюкопиранозных остатков: 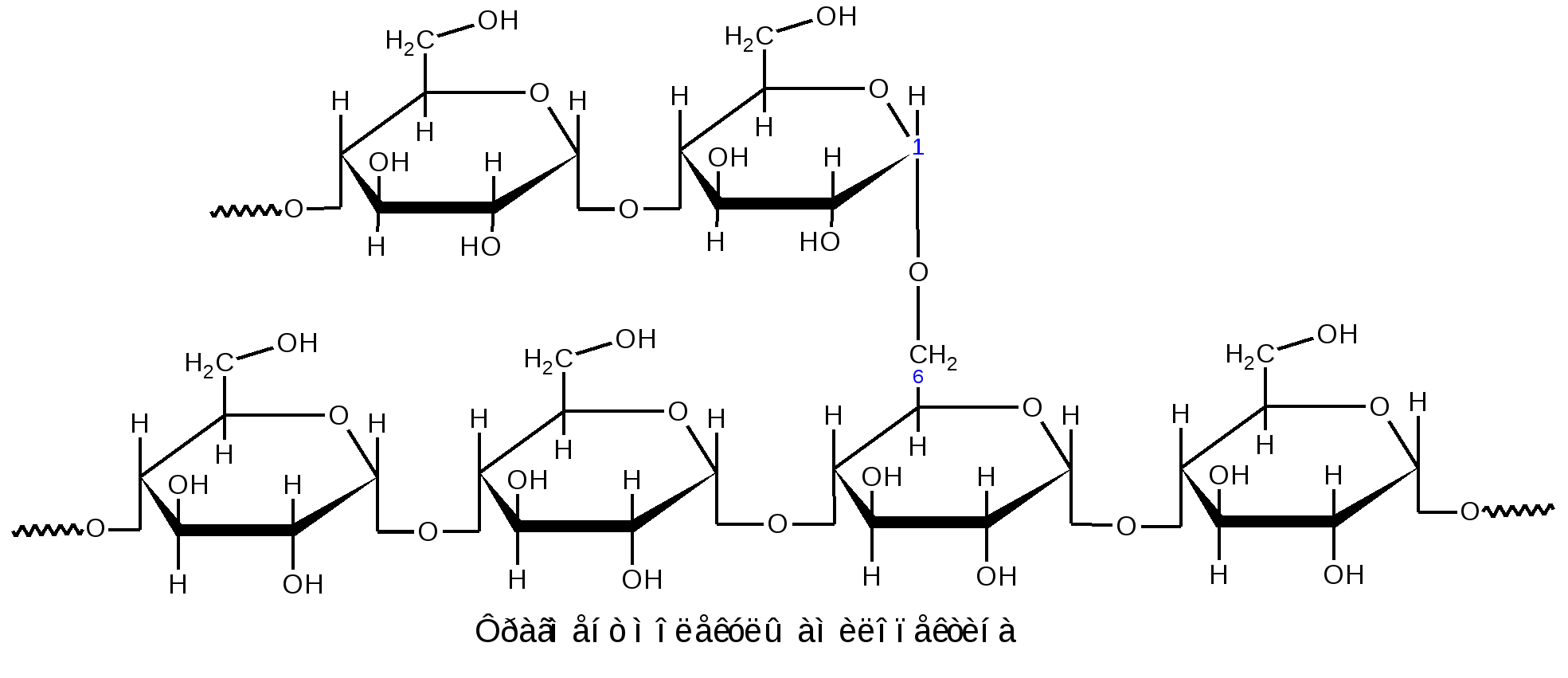 Амилопектин дает с йодом фиолетовое окрашивание; он расщепляется -амилазой на 50–60 %, причем наряду с мальтозой всегда образуются остаточные декстрины. Амилопектин сильно набухает в воде. При приготовлении клейстера из природного крахмала амилопектин препятствует ретроградации амилозы. Молекулярная масса амилопектина значительно выше, чем у амилоз и достигает 1–10 млн. Гликоген, или животный крахмал, является структурным и функцио-нальным аналогом растительного крахмала (резервный полисахарид). Он содержится во всех клетках животного организма. Наиболее богаты гликогеном печень (до 10–20 %) и мышечная ткань (до 4 %). Он содержится также в некоторых низших растениях, например в дрожжах и грибах; крахмал некоторых высших растений по свойствам близок к гликогену. Гликоген можно выделить из животных тканей путем обработки их 5–10 % трихлоруксусной кислотой на холоде с последующим осаждением извлеченного гликогена спиртом. Гликоген – белый аморфный порошок, хорошо растворяющийся в воде. Растворы гликогена дают с йодом окрашивание от красно-бурого до красно-фиолетового цвета (отличие от крахмала). Он легко гидролизуется кислотами и ферментами (амилазами), давая в качестве промежуточных продуктов декстрины и мальтозу и при полном гидролизе – глюкозу. Молекулы гликогена построены по тому же типу, что и молекулы амилопектина (остатки -D-глюкопираноз соединены -1,4- и -1,6- гликозидными связями), но в отличие от амилопектина гликоген имеет еще большее разветвление цепей. Точки ветвления у него в центральных частях молекулы разделены лишь тремя- четырьмя глюкозными остатками; периферические цепи гликогена состоят в среднем из семи- десяти глюкозных остатков. Сильное разветвление способствует выполнению гликогеном энергетической функции, так как только при наличии большого числа концевых остатков можно обеспечить быстрое отщепление нужного количества молекул глюкозы. Менее разветвленное строение амилопектина связано с тем, что в растениях значительно медленнее протекают метаболические процессы и не требуется быстрый приток энергии, как это иногда бывает необходимо животному организму (стрессовые ситуации, физическое или умственное напряжение). Молекулярная масса гликогена велика, она исчисляется миллионами. Такой размер молекул содействует выполнению функции резервного углевода. Так, макромолекула гликогена из-за большого размера не проходит через мембрану и остается внутри клетки, пока не возникнет потребность в энергии. Целлюлоза, или клетчатка – наиболее распространенный в растительном мире полисахарид. Она является главной составной частью клеточных стенок растений. Целлюлоза придает растительной ткани механическую прочность и эластичность, исполняя роль опорного материала растений. В природе целлюлоза не встречается в чистом виде. Волокна хлопка содержат 96–98 % целлюлозы, в различных видах древесины содержание ее составляет 40–60 %. Волокна льна и конопли состоят преимущественно из клетчатки. Важнейшими спутниками целлюлозы являются лигнин, гемицеллюлозы, пектиновые вещества, смолы и жиры. Способы отделения целлюлозы от спутников основываются на ее высокой стойкости к различным реагентам. Целлюлозу обычно выделяют, обрабатывая растительные материалы (хлопок, древесную щепу) растворами щелочи (натронный и сульфатный способы) или бисульфита кальция (сульфитный способ) под давлением при температуре от 120–160 С. Спутники целлюлозы при такой обработке растворяются. Попытки химического синтеза целлюлозы пока безуспешны. Наиболее чистыми образцами клетчатки, с которыми приходится часто встречаться, является вата, полученная из хлопка, и фильтровальная бумага. Поскольку целлюлоза является важным сырьем для получения химических волокон, пленок, пластмасс, лаков, бумаги и т. п., производство ее осуществляется в огромных масштабах. Структурной единицей целлюлозы является -D-глюкопираноза, звенья которой связаны -1,4-гликозидными связями. Это подтверждается тем, что при частичном гидролизе клетчатки образуется дисахарид целлобиоза, имеющий тоже -1,4-гликозидную связь. Строение клетчатки можно выразить следующей формулой: 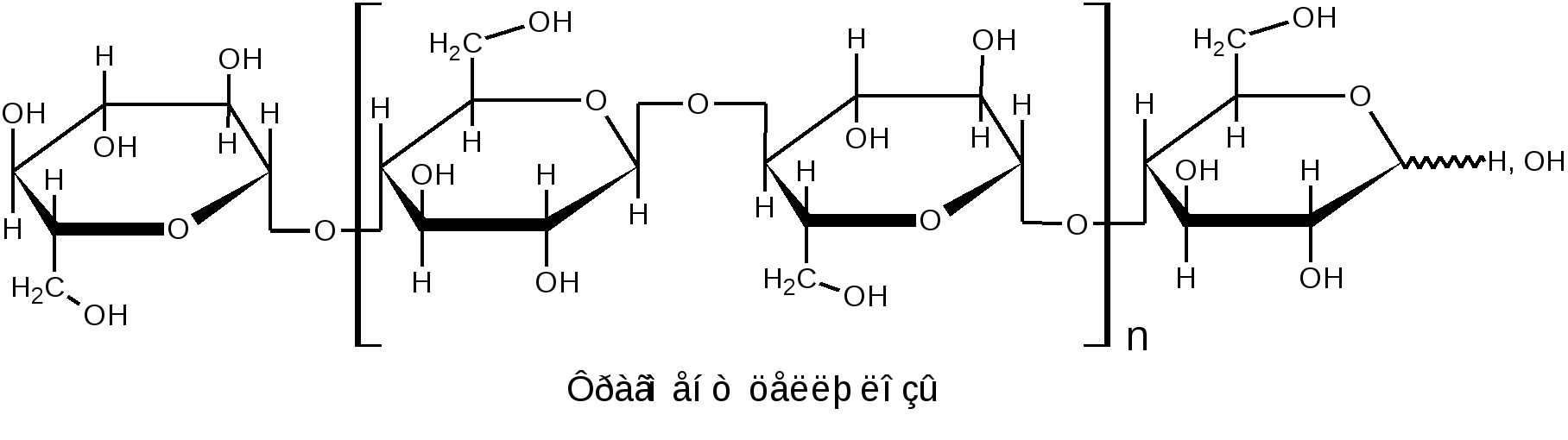 Макромолекулярная цепь целлюлозы не имеет разветвлений, в ней содержится 2500–12000 глюкозных остатков, что соответствует молекулярной массе от 400 тыс. до 1–2 млн. Строение целлюлозы, учитывающее конформацию, можно представить в виде формулы:  -Конфигурация гликозидной связи приводит к тому, что макромолекула целлюлозы имеет строго линейное строение. Этому способствует образование водородных связей внутри цепи, а также между соседними цепями. Такая упаковка цепей обеспечивает высокую механическую прочность, волокнистость, нерастворимость в воде и химическую инертность, что делает целлюлозу прекрасным материалом для построения клеточных стенок растений. Целлюлоза не расщепляется обычными ферментами желудочно-кишечного тракта, но она является необходимым для нормального питания балластным веществом. Сравнивая строение клетчатки со строением амилозы, мы видим, что различие сводится только к другому расположению кислородных мостиков, соединяющих глюкозидные остатки (амилоза имеет -1,4-, а целлюлоза -1,4-гликозидные связи). Однако эта, казалось бы, незначительная деталь строения приводит к огромным различиям в свойствах: в то время как амилоза относится к питательным веществам и очень легко расщепляется, целлюлоза является веществом, крайне стойким как к механическим, так и к химическим воздействиям. По внешнему виду целлюлоза – аморфное вещество. Однако рентгенографические исследования указывают на значительную упорядоченность (кристалличность) её структуры. Целлюлоза растворима в ограниченном числе растворителей, к ним относятся: водный раствор медноаммиачного комплекса [Cu(NH3)4](OH)2 – реактив Швейцера; водные растворы некоторых органических четвертичных оснований, например, гидроксид диметилдифениламмония; концентрированные минеральные кислоты (фосфорная, соляная, серная); трифторуксусная кислота; некоторые апротонные растворители; растворы хлорида цинка и некоторых других солей. Как и другие высшие полисахариды, целлюлоза не обладает восстанавливающей способностью. Целлюлозе, как и глюкозе, свойственны реакции многоатомного спирта, т. к. каждый элементарный остаток C6H10O5 содержит три гидроксила. В связи с этим формулу целлюлозы часто сокращённо пишут в виде [C6H7O2(OH)3]n. При нагревании целлюлоза разлагается, не плавясь. В присутствии кислорода заметное разложение наступает уже при температуре 110 оС. Без доступа кислорода разложение начинается при 140–150 оС. Выше 260 оС происходит бурное разложение целлюлозы с выделением сложной смеси летучих органических соединений и воды. Все важнейшие реакции целлюлозы связаны либо с расщеплением её макромолекулы, например, при действии гидролизующих агентов или окислителей, либо с получением производных целлюлозы как многоатомного спирта (реакции по ОН-группам или реакции с заменой ОН-групп другими функциональными группами). Гидролиз целлюлозы происходит под влиянием кислотных катализаторов, которые сильно ускоряют гидролиз гликозидных связей, весьма стойких к действию щелочей. Последовательность стадий гидролитического распада целлюлозы выражается следующей схемой: Целлюлоза Гидроцеллюлоза Целлодекстрины Целлоолигосахара Целлобиоза D-глюкоза Чтобы провести полный гидролиз клетчатки до глюкозы, её сначала растворяют в серной кислоте, разбавляют раствор водой и кипятят определённое время: H2SO4 (C6H10O5)n + n H2O n C6H12O6. Этот процесс имеет важное значение в технике, он служит основой производства этилового спирта гидролизом древесины с последующим сбраживанием образовавшейся при гидролизе глюкозы: C6H12O6 2 C2H5OН + 2 СO2. При действии окислителей (озон, перекись водорода, хлорноватистая кислота и её соли, разбавленная азотная кислота, перманганат калия и т.п.) получаются сложные смеси продуктов окисления, называемые оксицеллюлозами. При окислении механическая прочность целлюлозных материалов падает. Характерные химические изменения состоят в повышении содержания карбонильных и карбоксильных групп и в более или менее интенсивном разрыве полимерных цепей. Важное практическое значение имеет реакция взаимодействия целлюлозы с растворами NаОН (мерсеризация), приводящая к образованию так называемой алкалицеллюлозы, или щелочной целлюлозы. Взаимодействие целлюлозы с концентрированными водными растворами щелочей приводит преимущественно к образованию молекулярного соединения, но возможно также и образование некоторого количества алкоголятов целлюлозы. Процесс получения алкалицеллюлозы может быть описан следующей схемой: [C6H7O2(OH)3]n + mn NaOH [C6H7O2(OH)3 m NaOH]n Процесс мерсеризации целлюлозы сопровождается сильным ее набуханием. Алкалицеллюлоза при действии воды распадается снова на целлюлозу и едкую щелочь. Регeнерированнная целлюлоза (гидрат-целлюлоза) имеет уже другую физическую структуру. Мерсеризация хлопчатобумажных тканей широко используется для придания им лучшей окрашиваемости и более красивого внешнего вида. Щелочная целлюлоза имеет также большое значение в технике как промежуточный продукт в производстве вискозного волокна и эфиров целлюлозы. Целлюлоза, как и любой другой спирт, образует простые и сложные эфиры. В зависимости от условий проведения реакции можно получить производные по одному, двум или всем трем гидроксилам. Максимальная степень этерификации отвечает триэфиру целлюлозы. О +NaOH дним из основных методов получения простых эфиров целлюлозы является действие на щелочную целлюлозу алкилгалогенидов или диалкилсульфатов в присутствии щелочи: [C6H7O2(OH)3 m NaOH ]n + nm C2H5Cl [C6H7O2(OH)3-m (OC2H5)m]n + m n NaCl + m n H2O +NaOH [C6H7O2(OH)3 m NaOH ]n + nm (CH3)2SO4 [C6H7O2(OH)3-m (OCH3)m]n + m n CH3NaSO4 + m n H2O Из простых эфиров целлюлозы наибольшее применение в технике имеют метил-, этил- и бензилцеллюлозы. При метилировании целлюлоза приобретает некоторую растворимость в воде. Метилцеллюлоза используется в основном как загуститель (вместо крахмала) в текстильной, косметической и пищевой промышленности. Этилцеллюлоза обладает высокой механической прочностью, термо- и морозостойкостью. Она используется в производстве пластмасс, пленок и лаков. Среди сложных эфиров целлюлозы важнейшими являются азотнокислые, уксуснокислые и ксантогенат целлюлозы. Азотнокислые эфиры (нитраты) целлюлозы обычно получают действием на целлюлозу смеси азотной и серной кислот, H2SO4 [C6H7O2 (OH)3]n +3n HNO3 [C6H7O2(ONO2)3 ]n+ 3nH2O причем серная кислота служит водоотнимающим средством. Общим свойством нитратов целлюлозы является их чрезвычайная горючесть и огнеопасность. Продукт полной этерификации – тринитрат целлюлозы должен содержать в соответствии с формулой 14,1 % азота. На практике получается продукт с несколько меньшим содержанием азота (12,5–13,5 %), называемый в технике «пироксилин». Пироксилин применяется в производстве бездымного пороха, где его желатинируют смесью органических растворителей (ацетон и уксусноамиловый эфир), в некоторых случаях с добавлением нитроглицерина. Продукт нитрации клетчатки, содержащий 10 % азота, в технике известен под названием «коллоксилин». При действии на него смеси спирта и эфира образуется вязкий раствор, так называемый коллодий. Коллодий- густая, клейкая жидкость, оставляющая при высыхании блестящую пленку. Нитрат целлюлозы в смеси с камфорой образует пластмассу – целлулоид, имевший раньше значительное применение для изготовления изделий широкого потребления. Теперь целлулоид вытеснили гораздо более пожаробезопасные материалы, в частности, ацетаты целлюлозы. Уксуснокислые эфиры (ацетаты) целлюлозы получают действием на клетчатку уксусным ангидридом в присутствии уксусной кислоты и небольших количеств серной кислоты как катализатора: [C6H7O2 (OH)3]n +3n (CH3CO)2O [C6H7O2(OCOCH3)3 ]n+ 3n CH3COOH. Триацетилцеллюлозу называют первичным ацетатом. Наибольшее практическое значение имеет вторичный ацетат, близкий по содержанию ацетильных групп к диацетату и получаемый частичным гидролизом первичного ацетата: [C6H7O2(OCOCH3)3 ]n + n H2O [C6H7O2 (OH) (OCOCH3)2]n + n CH3COOH. Ацетат целлюлозы является ценным материалом для производства негорючей кинопленки, пластмасс – этролов и особенно ацетатного шелка. Для производства ацетатного шелка растворы ацетата целлюлозы в ацетоне продавливают через фильеры-колпачки с тонкими отверстиями; далее ацетон в токе нагретого воздуха испаряется, и струйки раствора превращаются в тончайшие нити из ацетата целлюлозы. К |