Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА XII |
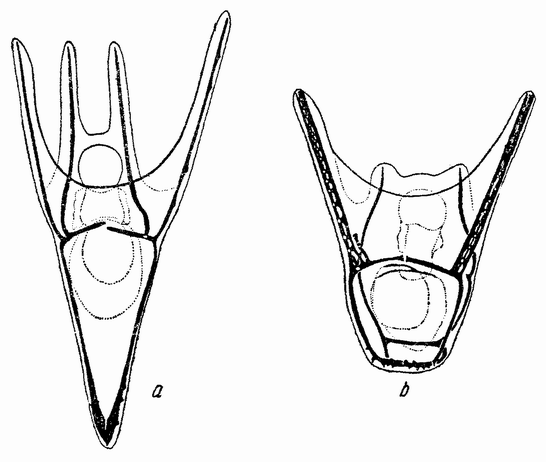 |
| Рис. 82. а — плутеус Strongylocentrotus; b — плутеус Sphaerechinus (по Гербсту). |
совершаются теперь с помощью системы трубчатых ножек, располагающихся вдоль пяти лучей вновь образованного тела. Личинка имела двустороннюю симметрию, взрослое же животное радиально симметрично.
Морские кольчатые черви и моллюски проходят свободноплавающую стадию, известную под именем трохофоры (рис. 83 а). Трохофора имеет два ряда ресничек, а на своем переднем конце несет группу ресничек, в основании которых находится очень примитивный нервный центр. Пищеварительный тракт состоит из рта, пищевода, желудка, {143} кишечника и заднепроходного отверстия. Между наружной стенкой пищеварительного тракта и внутренней стенкой тела имеется большое пространство с немногочисленными мускульными клетками, тянущимися вокруг и вдоль стенок. У кольчатых червей на заднем конце имеется небольшая группа мезодермальных клеток, из которых развиваются будущие сегменты и мускулатура. Превращение трохофоры в червя происходит постепенно. Задний конец трохофоры вытягивается (рис. 83 b); из мезо-
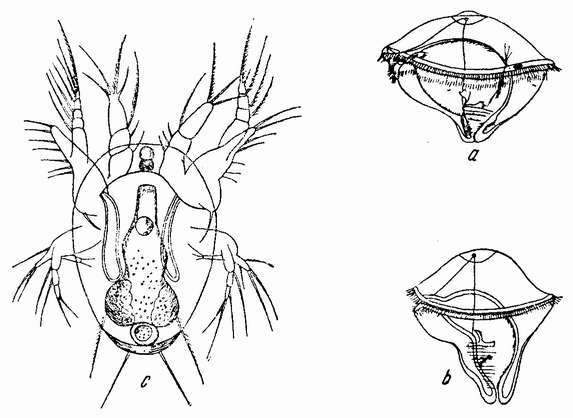 |
| Рис. 83. а — трохофорная личинка кольчатых червей; b — более поздняя стадия ее, обнаруживающая удлинение заднего конца и развитие сегментов: с — личинка ракообразных — науплиус (по Гатчеку из Коршельта и Гейдера). |
дермы одна за другой образуются парные группы клеток. Каждая пара содержит полость, внутренние стенки которой образуют мускулатуру пищеварительного тракта, а наружные — мускулатуру стенки тела. Между располагающимися друг за другом сегментами образуется септа; головной конец редуцируется; мозг увеличивается, и эктодермальное утолщение вдоль вентральной стороны становится нервной струной.
Превращение трохофоры моллюсков происходит несколько отличным путем. В качестве выделения кожного {144} покрова на дорзальной стороне развивается раковина, которая впоследствии облекает животное, когда оно сокращается. Утолщение вентральной стенки образует мускулистую ногу. Взрослое животное не разделено на
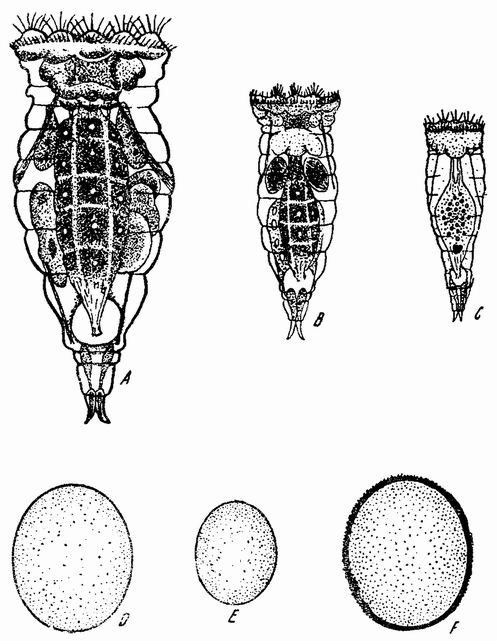 |
| Рис. 84. Hydatina senta. A — взрослая самка; В — молодая самка вскоре после вылупления; С — взрослый самец; D — партеногенетическое яйцо; Е — яйцо, дающее самца; F — покоящееся яйцо (по Уитнею). |
членики, как аннелиды, а состоит из одного единственного переднего сегмента или во всяком случае имеет не более одного добавочного. Взрослые моллюск и аннелида эволюционировали совершенно различными путями.
Наличие у обеих групп общего типа личинки интерпретировалось как доказательство того, что эта личинка {145} является предком, из которого развились обе группы. Если принять в расчет другие соображения, то кажется более вероятным, что присутствие у обеих групп трохофоры скорее означает, что они произошли от общего предка, который проходил через сохранившуюся до сих пор стадию трохофоры.
Идентичность их типов дробления подтверждает этот взгляд, так как стадии дробления конечно не были взрослым предком.
Существует группа пресноводных и морских животных, так называемые коловратки (рис. 84 а), которые во многих отношениях напоминают трохофорную личинку (рис. 83). Развитие коловраток никогда не идет дальше этой стадии, но у них образуется пара яичников или семенников. Были высказаны предположения, что коловратки представляют собой задержавшуюся трохофорную стадию какой-то древней аннелиды (или моллюска), которая больше не достигает прежнего взрослого состояния.
Большинство низших ракообразных имеет маленькую свободноплавающую личинку с шестью конечностями, (рис. 83 с), называемую науплиусом, который одно время тоже считался предком, из которого развились все ракообразные. Имеется, однако, больше оснований рассматривать науплиус как сохранившуюся до наших дней личинку предков ракообразных.
Большинство низших насекомых проходит через последовательный ряд стадий, каждая из которых более совершенна, чем предыдущая. Каждая стадия заканчивается линькой, при которой сбрасывается наружная кутикула, мешающая дальнейшему росту. Тем временем под старой кутикулой развивается новый мягкий покров. Заметное увеличение величины личинки происходит немедленно, прежде чем новая кутикула затвердевает. У наиболее высокоорганизованных групп насекомых — жуков, бабочек, мух, пчел и муравьев — имеется характерная личиночная стадия, сильно отличающаяся от взрослой формы. Организм на этой стадии называется личинкой или гусеницей. Переход от личинки к взрослой стадии у этих групп происходит внезапно. Совершается типичный метаморфоз. Между личинкой и взрослой формой находится неподвижная стадия, так называемая куколка, у которой внутри пуппария из дисков недиференцированных клеток, так называемых имагинальных дисков, формируются органы взрослого {146} животного. Большинство личиночных органов при этом полностью теряется и рассасывается.
Не пытаясь описывать метаморфоз насекомых, будет достаточным указать, что типы личинок этих высших групп не могут считаться полностью соответствующими личинкам предков, хотя некоторые признаки последних и могли быть сохранены. Личинки являются сами по себе высоко приспособленными формами, которые, может быть, настолько отошли от исходного типа, насколько сами взрослые формы отошли от других членов прародительской группы. Например, гусеница бабочки грызет свою пищу с помощью роговых жвал, тогда как сама бабочка имеет трубку для всасывания нектара из растений. У гусеницы развивается новый комплект конечностей; сложные глаза у нее отсутствуют. Личинка и взрослый организм стали настолько специализированными, что постепенный переход одного в другое невозможен. Поэтому, когда гусеница достигает своей конечной стадии развития, она перестает двигаться, вьет вокруг себя кокон и из ее имагинальных дисков недиференцированных клеток образуются органы бабочки. Сказать точно, каким образом это приспособление произошло исторически, — трудно, но у некоторых других групп имеются переходные стадии, которые делают эту трудность преодолимой.
У позвоночных нет вполне типичной для всех них личиночной стадии, но у амфибий таким типом личинки является головастик лягушки. Он тоже, подобно гусенице, представляет сильно специализированную личинку и в конце личиночного периода претерпевает внезапный метаморфоз.
Головастик лягушки многими своими особенностями напоминает соответствующую стадию более простых групп амфибий — тритонов и саламандр — и так как личинки последних по всем своим признакам, за исключением величины, очень похожи на взрослые стадии, то может показаться, что головастик представляет собой взрослую саламандру, повторяющуюся в развитии лягушки. Это и будет приложением так называемого биогенетического закона Геккеля. Однако имеется много оснований для того, чтобы счесть последнюю интерпретацию ошибочной. Головастик не является рекапитулировавшей стадией саламандры, а стадией развития общей и для лягушки, и для саламандры.
Когда головастик вырастает до своих полных размеров, {147} что у различных видов может продолжаться от нескольких недель до одного года или больше, он прекращает питаться и в течение нескольких дней претерпевает ретрогрессивные изменения в одних из своих органов и прогрессивные в других. Хвост головастика укорачивается и его внутренние ткани рассасываются. Они пожираются фагоцитами или белыми кровяными клетками, которые несут продукты их распада назад в организм. Большая часть жаберного участка рассасывается. Пищеварительный тракт головастика, представляющий собой длинную свернутую трубку, во время метаморфоза укорачивается и стенки его утолщаются. Роговые зубы во рту исчезают и рот расширяется. Жаберные щели закрываются, а сами жабры рассасываются. Кровеносные сосуды, проходящие через жаберные дуги, частично рассасываются, частично же превращаются в некоторые сосуды взрослого животного. Легкие, которые развиваются к тому времени в виде пары мешковидных выростов пищевода, начинают функционировать, как только в них набирается через нос воздух. Происходят дальнейшие видимые прогрессивные изменения. Передние и задние конечности, присутствующие у головастика в виде небольших рудиментарных органов, внезапно начинают увеличиваться. До начала метаморфоза передние конечности очень малы, во время же него они быстро растут и пробиваются сквозь закрывающую их до этого времени складку. Задние конечности, которые длиннее передних, тоже быстро растут. Интересно отметить, что оба обратных процесса происходят одновременно: одни органы дегенерируют, тогда как другие растут.
Метаморфоз тесно связан с изменениями в щитовидной железе, являющейся органом внутренней секреции, вырабатывающим гормон (Thyroxin), т. е. вещество, абсорбирующееся из железы кровью и вызывающее некоторые из вышеописанных изменений в теле. При содержании головастика в воде с примесью йодистых соединений или путем инъекции сходных веществ переход к лягушке может совершиться задолго до обычного срока.
Одна из хвостатых амфибий, а именно мексиканский аксолотль, достигает половой зрелости, когда у нее еще присутствуют жаберные щели и жабры. Аксолотль разводится обычно в этом состоянии. Его взрослая форма — сухопутная саламандра — развивается только при исключительных условиях. Созревание и функционирование {148} половых клеток — яиц и спермиев — в то время, когда особь находится еще на стадии личинки, называется неотенией. Можно подозревать, что это явление имеется и у других представителей хвостатых амфибий, а также и у некоторых других групп животных. Например, асцидии проходят через свободноплавающую личиночную стадию (рис. 85 а), напоминающую головастика лягушки. Существует также
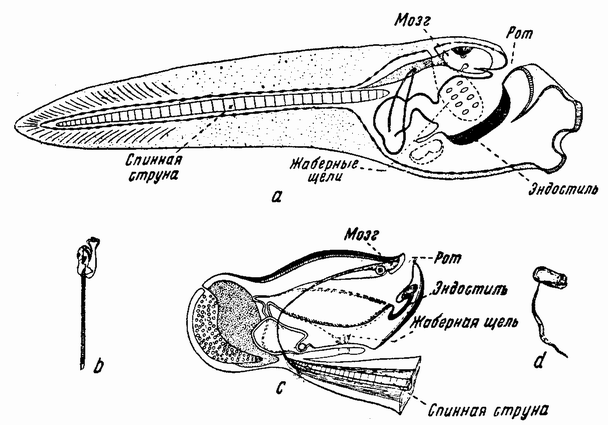 |
| Рис. 85. а, b — стадия головастика у асцидии; с, d — свободноплавающая аппендикулярия. |
группа свободноплавающих пелагических асцидии, Append icularia (рис. 85, с), имеющая основные черты стадии головастика других асцидии. Эти аппендикулярии размножаются, но никогда не превращаются в форму, характерную для других членов этой группы. Они, вероятно, являются неотеническими формами.
В противоположность животным, обладающим многочисленными мелкими яйцами, дающими начало свободноживущим личинкам, существуют другие животные, образующие небольшое число крупных яиц, содержащих достаточно пищевых веществ для поддержания жизни зародыша в течение его последующего развития до той стадии, {149} которая всеми своими признаками, кроме размеров, напоминает взрослый организм. В этом случае можно сказать, что развитие является прямым. К зародышам с прямым развитием, которые до относительно поздней стадии своего развития находятся внутри материнского организма, применяется термин плод. Плод прикреплен к матке особым органом зародыша — плацентой, которая так соединяет кровеносную систему зародыша с кровеносной системой материнского организма, что зародыш абсорбирует из нее питательные вещества.
В большинстве крупных групп имеются виды с прямым развитием и эти виды обычно имеют очень крупные яйца.
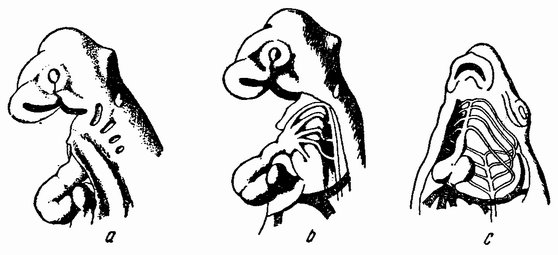 |
| Рис. 86. а — голова цыпленка, обнаруживающая четыре жаберных щели (вид сбоку); b — дуги аорты; с — сердце и дуги аорты у рыбы (по Гессе). |
«Желток» куриного яйца в двадцать семь миллионов раз превышает объем яйца ланцетника. Наилучше изученные и наиболее интересные случаи известны у позвоночных. Огромные яйца имеют ящерицы и птицы, и молодь у них при вылуплении или при рождении имеет форму взрослого организма. Развитие млекопитающих весьма похоже на развитие птиц и ящериц, несмотря на тот факт, что оно берет начало из очень маленького яйца.
Сравнение развития этих типов друг с другом и с более низко организованными членами группы обнаруживает некоторые интересные отношения. Во-первых, наличие У ящериц, птиц и позвоночных (рис. 86 а, b) таких систем органов, как жаберные щели, которые напоминают эти же органы у свободноживущих личинок рыб (рис. 87) и амфибий; {150} во-вторых, присутствие эмбриональных новообразований, приспосабливающих зародыш к новым условиям, как, например, амнион и аллантоис (рис. 31). Кроме того, имеется много второстепенных признаков, относящихся к одной из этих двух категорий. Сравнение приводит нас назад к стадиям развития рыбы и амфибии. Рыбы, как хрящевые (акулы), так и костистые (Teleosti), дышат с помощью жаберной системы (рис. 86 и 87), состоящей из расположенных по бокам шеи жаберных щелей, которые внутри открываются в пищевод, а снаружи в воду. Между жаберными щелями находятся жаберные дуги, по которым кровь проходит от вентрального сердца к дорзальной аорте. На дугах име-
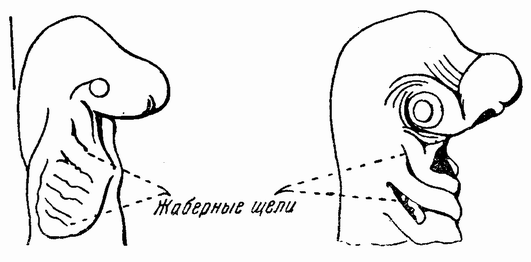 |
| Рис. 87. Голова молодой хрящевой рыбы сбоку; видны жаберные щели (по Седжвику). |
ется множество нитевидных жабер, содержащих капиллярные кровеносные сосуды. В жабрах кровь окисляется. Акт дыхания состоит в набирании в рот воды и в пропускании ее сквозь жаберные щели над жабрами. Эта система появляется в развитии рыб очень рано, независимо от того, происходят ли они из крупных яиц, как у акул, или из мелких, как у костистых рыб. Система эта, однако, не функционирует до вылупления молодого животного из яйца.
У эмбрионов ящериц, птиц и млекопитающих (рис. 88) на соответствующей стадии тоже появляются жаберные щели. Но они никогда не функционируют как органы дыхания, а скоро закрываются. На других жабры не образуются, но кровеносные сосуды, несущие кровь через дуги, функционируют с самого начала, неся кровь от сердца к спинной аорте. На более поздних стадиях некоторые из этих кровеносных {151} сосудов исчезают, другие же сохраняются, становясь частями кровеносной системы взрослого организма. Важными моментами являются при этом, во-первых, то, что у зародышей более высоко организованных групп дыхательный аппарат появляется на той же стадии, что и у более низко организованных групп; во-вторых, что у зародыша он выполняет функцию переносчика крови от сердца к спинной стороне; и в-третьих, что аортные дуги представляют собой структуру, из которой возникают более специализированные системы взрослого организма.
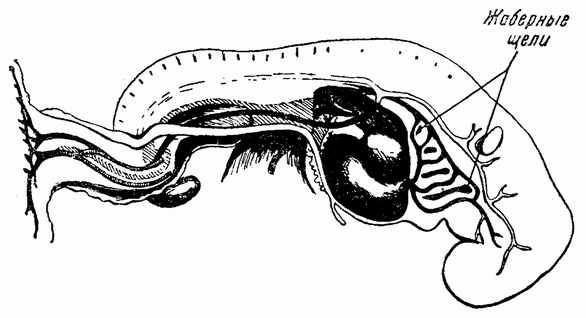 |
| Рис. 88. Человеческий эмбрион; видны сердце, дуги аорты и жаберные щели (по Гису из Маршаля). |
Амфибии представляют собой интересное промежуточное состояние. Они ближе, чем рыбы, к более высоко организованным типам, хотя они могут и не быть в числе их непосредственных потомков. Более того, некоторые амфибии являются одними из самых первых позвоночных животных, которые покинули воду, для того чтобы жить на суше, и которые обнаруживают переход от жабер к легким. Лягушки и многие хвостатые амфибии, живущие в настоящее время на суше, уходят для откладки яиц в воду, где протекает развитие их молодых стадий. Молодь сначала дышит при помощи эмбриональных наружных жабер, которые замещаются внутренними жабрами, подобными таковым у рыб. Жаберная система рассасывается, когда выросшие личинки покидают воду для сухопутной жизни, причем это рассасывание и трансформация в основном происходят так же, как и у птиц и млекопитающих. {152} Таким образом, ранние стадии молодых лягушек протекают так же, как и у воднодышащих рыб, а их взрослые стадии подобны таковым рептилий, птиц и млекопитающих.
Сходство молодых стадий высших групп позвоночных было подмечено и истолковано еще более ста лет назад, т. е. уже в то время, когда теория органической эволюции еще не была распространена. Этому сходству тогда было дано только формальное объяснение. Наиболее известное положение, именуемое в настоящее время законом Бэра, было установлено в 1828 году. Положение Бэра часто смешивается с биогенетическим законом Геккеля, который, однако, совершенно отличен и предполагает принятие эволюционной теории. Четыре закона Бэра таковы:
1. Наиболее общие признаки крупных групп появляются у эмбриона раньше, чем специализированные признаки.
2. За наиболее общими структурами возникают менее общие и так до тех пор, пока, наконец, не возникнут наиболее специализированные образования.
3. Эмбрион каждого данного взрослого животного не повторяет при своем развитии другие взрослые формы, а, наоброт, дивергирует с ними.
4. В основном эмбрион высшего животного никогда не бывает похож на взрослую стадию другой животной формы, а походит только на ее зародыш.
Так называемый биогенетический закон более определенен. Он гласит, что эмбрионы представителей высших групп проходят через взрослые стадии более низко организованных представителей этой же группы, которые как бы конденсированы в их развитии. Это положение так картинно изображает некоторые особенности развития, что оно стало одной из наиболее популярных теорий биологии. Его большая простота скрывает его запутанность и создает впечатление «объяснения», в то время как оно скорее служит для сокрытия исторического родства. Теперь, когда мы лучше понимаем, каким образом появляются новые наследственные изменения, мы находим, что нет постоянной связи между исходным признаком и появлением нового, как это подразумевается в биогенетическом «законе». Во многих случаях, может быть, даже в большинстве, новый конечный признак просто замещает исходный. Эмбрион развивает новый признак не после того, как он проходит через последнюю стадию исходного признака, хотя иногда это и может случиться, а новый признак занимает место {153} исходного. Так, например, мутант дрозофилы, имеющий рудиментарные крылья, при своем развитии не проходит через стадию с большими крыловыми пластинками или крупными дисками, а крылья непосредственно развиваются у него из дисков пропорционально уменьшенной величины. То же имеет место почти для всех известных мутантных форм, причем это верно не только для тех случаев, в которых новый орган оказывается уменьшенным, как в случае зачаточных крыльев, но и когда новый признак является добавлением чего-либо лишнего к тому, что было до этого.
Правда, имеются некоторые случаи, в которых новый признак налагается на старый, причем последний может до некоторой степени сохраняться. Например, некоторые жуки имеют черные пятна на желтом фоне. Известны черные мутанты, развитие которых проходит стадию, на которой присутствуют пятна на светлом фоне, но последний скоро темнеет, закрывая собой пятна. В этом случае можно сказать, что новый признак добавляется к старому в конце ряда, здесь, может быть, будет правильным сделать вывод, что взрослая стадия предка сохранилась в развитии последующей формы. Однако такие случаи являются исключениями. Правило же, как было установлено выше, состоит в том, что первоначальная конечная стадия выпадает из развития и замещается новым признаком.
Хотя Геккель и считал, что эмбриональные стадии по большей части подобны взрослым стадиям предков, но он был знаком также и с эмбриональными приспособлениями, которые за взрослые стадии никак не могут быть сочтены; таковы, например, аллантоис, амнион и желточный мешок птиц и млекопитающих. Он называл их фальсификацией филогенетически предшествовавших стадий. Они являются эмбриональными приспособлениями. Вскоре были найдены основания для скептического отношения к толкованию многих личиночных и эмбриональных стадий как предков ныне живущих форм. Таковы, например, науплиус ракообразных и трохофора кольчатых червей, которые рассматриваются теперь только как специализированные личиночные формы, и широкое распространение их в группе не есть достаточное доказательство того, что эти личинки соответствуют взрослым стадиям предков. Другими словами, если теория рекапитуляции является «законом», то он имеет так много исключений, что становится бесполезным и часто ошибочным. Более того, как было {154} установлено выше, он предполагает такой способ «прибавления» новых признаков, который не соответствует многочисленным фактам, которыми мы располагаем.
Теория гена с ее допущениями полностью согласуется со всем тем, что известно относительно развития вновь появляющихся признаков. Мутационное изменение в гене может быть таким, что его эффект становится видным только по достижении последней стадии развития, причем это изменение может эту стадию полностью изменить. Опять-таки может быть такого рода генное изменение, которое, влияя на личиночную стадию, изменяет один или большее число из ее признаков. Такое изменение может либо затрагивать взрослые стадии, либо нет. A priori должно казаться, что чем раньше совершается изменение в принятом типе развития, тем более вероятно, что оно будет затрагивать также и поздние стадии. Вся цепь явлений развития так сложна, что, вероятно, много труднее получить изменение в ранней стадии, не вызывая нарушения последних стадий, чем наоборот. Это обстоятельство могло создать благоприятные условия для сохранения ранних эмбриональных и личиночных стадий крупных групп.
Ни все законы Бэра, ни биогенетический закон Геккеля не находятся в соответствии с известными в настоящее время фактами изменчивости и развития. Возможно дать объяснение, много более простое и более согласующееся с этими фактами. Жаберные щели млекопитающих должны сравниваться с таковыми не у взрослых рыб, а с жаберными щелями молодых организмов на той же стадии. По мере эволюирования высших позвоночных из низших они сохранили много признаков начальных стадий своих предков. При желании можно было бы, став на телеологическую точку зрения, сказать, что либо этот путь является единственно «известным» для них методом достижения взрослой стадии, либо же, иначе, что они нашли, что наиболее простым способом для развития более сложных структур является пристройка их на тех, которыми организм пользовался уже миллионы лет. Такая точка зрения может быть названа принципом эмбрионального консерватизма. Выражаясь менее антропоморфично, этот принцип, которому не обязательно давать громкое название закона, а который может именоваться лишь принципом эмбрионального консерватизма, заключается в том, что высшие и низшие члены группы проходят одни и те же стадии развития.
| << | | {155} | | >> |