Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА XI |
| Cidaris | Часы | Lytechinus | Часы |
| Бластула (плавающая) | 16—18 | Бластула (плавающая) | 5,5 |
| Гаструла (начало образования) | 20—23 | Мезенхима | 8 |
| Мезенхима | 23—26 | Гаструла (начало образования) | 9 |
| Хроматофоры | 44 | Хроматофоры | 15—16 |
| Скелет (начало образования) | 72—73 | Скелет (начало образования) | 15—16 |
| Плутеус | 120 | Плутеус | 24 |
При оплодотворении яиц Cidaris спермой Lytechinus скорость дробления равна таковой у Cidaris; бластула тоже подобна бластуле Cidaris. У Cidaris мезенхимные клетки развиваются из внутреннего конца первичного кишечника, тогда как у Lytechinus они начинают мигрировать внутрь до начала гаструляции. Мезенхимные клетки гибрида возникают по краям и вокруг основания первичного кишечника, а не из внутреннего конца, как у Cidaris. Представляется поэтому, что на место возникновения мезенхимных клеток влияет сперматозоид Lytechinus.
Форма молодой треугольной личинки, получающейся при оплодотворении яиц Sphaerechirius сперматозоидами Strongylocentrotus, подобна таковой материнского вида, но на стадии плутеуса, которая следует непосредственно за стадией треугольной личинки, становится видимым влияние отцовских хромосом. Имеются многочисленные описания более взрослых стадий плутеусов гибридов между различными видами морских ежей, и практически у всех из них могут быть обнаружены характерные черты обоих родительских типов. Немногочисленные попытки произвести генетический анализ признаков (таких, как форма игольчатого скелета, форма личинки, характер пигмента и т. д.) не были очень удачными, так как хотя и оказалось возможным установить доминирование одного признака над другим, но число участвующих факторов не может быть определено без получения еще одного поколения от гибридов. Только таким путем, {128} как и в большинстве случаев генетического анализа, из числовых отношений между вновь появляющимися типами может быть определено число участвующих в скрещивании факторов.
Общие выводы из этих данных ясны. Ранние стадии развития определяются протоплазмой яйца; влияние же хромосом сперматозоида сказывается, только позже. Это означает, что протоплазма яйца уже подверглась влиянию генов, содержащихся в самом яйце, и требуется время для того, чтобы отцовские гены вызвали изменение в протоплазме клеток эмбриона.
Опыты по гибридизации усложняются двумя другими моментами, именно влиянием среды и неправильностями в распределении хромосом, которые иногда имеют место на ранних стадиях дробления гибрида.
Действие температуры изучалось как на чистых, так и на гибридных плутеусах, причем некоторое влияние ее было обнаружено. У гибридов, повидимому, от температуры зависит большее или меньшее доминирование материнского или отцовского влияния. Щелочность или кислотность морской воды в некоторых случаях тоже, повидимому, влияет на результат.
В некоторых скрещиваниях было показано, что на первых или более поздних стадиях дробления яйца может иметь место элиминация единичных или даже нескольких хромосом. Так, при оплодотворении яиц Arbacia сперматозоидами Sphaerechinus могут быть элиминированы около 18 хромосом предположительно отцовского происхождения, что может объяснять преобладание материнских черт у гибридного плутеуса. В реципрокном скрещивании не происходит элиминации и тем не менее плутеус оказывается сходным с таковым материнской расы. Элиминация хромосом, нарушающая сбалансированный эффект генов, может иметь и дальнейшее влияние, вызывая у эмбриона патологические признаки.
Для гибридизации многократно использовались также костистые рыбы, отдаленные виды которых могут широко скрещиваться друг с другом, но только в некоторых скрещиваниях эмбрион развивается до взрослой стадии. Скорость дробления при этом равна скорости дробления у материнского вида за исключением случаев, когда она замедляется вследствие ненормального разделения хромосом. Но при скрещивании близко родственных видов было {129} установлено, что темп дробления может быть ускорен отцовскими генами, если отец принадлежал к типу с более быстрым дроблением. Развивающиеся пигментные клетки происходят не из пигмента, содержащегося в яйце, а пигмент развивается в них независимо и может быть подвержен влиянию отцовских генов. Элиминация хромосом описывалась в различных скрещиваниях. Многие эмбрионы погибают на ранних стадиях, что, вероятно, в одних случаях объясняется элиминацией и неправильными делениями хромосом, в других же это более вероятно объяснить несовместимостью эффектов генов обоих видов.
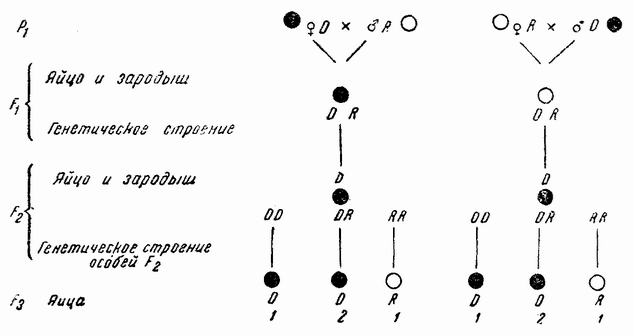 |
| Рис. 76. Схема материнского наследования у шелковичного червя. |
У молодых эмбрионов некоторых рас шелковичного червя пигмент появляется в эмбриональной мембране из веществ, предшествующих ему в яйце. При скрещивании рас с различно окрашенной эмбриональной мембраной цвет ее у потомства оказывается материнским. Когда из этих яиц были получены взрослые гибридные бабочки, то было найдено, что у их эмбрионов развился доминантный цвет независимо от того, был ли он привнесен отцом или матерью (рис. 76). Что здесь мы действительно имеем случай генной наследственности, было показано путем выведения еще одного поколения, в котором эмбрионы (F.) расщепились в отношении 3:1 (или более точно 1:2:1). Этот результат показывает, что эмбриональный признак {130} определяется генами, которые следуют менделевскому закону расщепления для единственной пары факторов (рис. 76). Другими словами, материнская наследственность принципиально не отличается от обычной менделевской. В этом случае предшественники пигмента уже заложены в яйце, благодаря влиянию генов в течение ранних стадий формирования яйца, и не сразу изменяются под действием отцовских генов, даже если последние доминантны. Но в этом случае эффект доминантных генов проявляется в следующем (F2) поколении. В этом поколении имеются три генетически различных класса особей; доминантные и
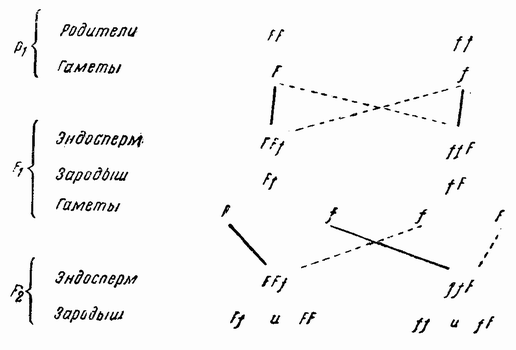 |
| Рис. 77. Схема образования эндосперма у кукурузы. |
рецессивные гены расщепились в F1 и каждый, действуя на развитие протоплазмы своего собственного яйца, образует в F3 тот или иной тип эмбриона.
У этой бабочки имеются некоторые расы, дающие две генерации в год, тогда как другие расы имеют только одну. Было показано, что это различие тоже обнаруживает материнское наследование и вне всякого сомнения обусловлено какой-то особенностью яйца или яйцевой мембраны, которая в свою очередь обусловлена генами.
Подобные же явления найдены у индийской кукурузы при образовании эндосперма семян. Эндосперм является соматической тканью, образующейся в зародышевом мешке — увеличенной клетке — путем соединения двух материнских {131} ядер и одного отцовского. Цвет эндосперма определяется доминантными и рецессивными генами, содержащимися в этих ядрах. Например: если в качестве материнского растения была взята мучнистая кукуруза (ff), а в качестве отцовского кремнистая, то эндосперм получится мучнистым (ff F), откуда следует, что две дозы или два гена мучнистости доминируют над одним геном кремнистости (рис. 77). Наоборот, если кремнистая кукуруза была материнским растением, а мучнистая — отцовским, то эндосперм получается кремнистым (FFf), так как два гена кремнистости доминируют над одним мучнистости. Правильность этих формулировок показана выведением следующих поколений, в которых оба признака эмбриона (F и f) расщеплялись как одна пара генов. Надо отметить, что в этих двух случаях протоплазма эндосперма не изменялась геном пыльцевого зерна, потому что два рецессива подавляют собой доминантный ген.
Наконец, интересный случай материнской наследственности был найден у некоторых пресноводных моллюсков (Lymnaea). Раковины этих моллюсков обычно закручены в правую или декстральную спираль, но случайно встречаются формы, имеющие левые или синистральные витки спирали. Все потомство данного выводка, включая гибридов, имеет либо декстральные, либо синистральные витки. Было также показано, что некоторые матери с синистральными витками дают только синистральные семьи, тогда как другие такие же самки — только декстральные. Наоборот, некоторые декстральные матери производят только синистральные семьи, в то время как другие могут давать декстральное потомство. Эти факты были сначала с генетической точки зрения очень загадочными, но теперь мы имеем для них удовлетворительное объяснение. Предположим, как на это указывают факты, что имеется доминантный ген для декстральных витков и рецессивный для синистральных и что гены эти содержатся в данной паре хромосом. Самооплодотворяющаяся декстральная улитка гетерозиготная по этим генам (Ll) образует при созревании два сорта яиц L и l. Подобным же образом будет два сорта сперматозоидов L и l. Самооплодотворение приведет к образованию трех генетически различных типов потомков — LL, Ll и ll, но все эти особи будут иметь декстральные завитки, потому что тип дробления был определен уже в яйце доминантным фактором L еще до выделения {132} полярных телец. Из этих трех типов первые два (LL и Ll) будут давать только декстральное потомство, а третий тип (ll), также имеющий декстральные витки, даст только сини-
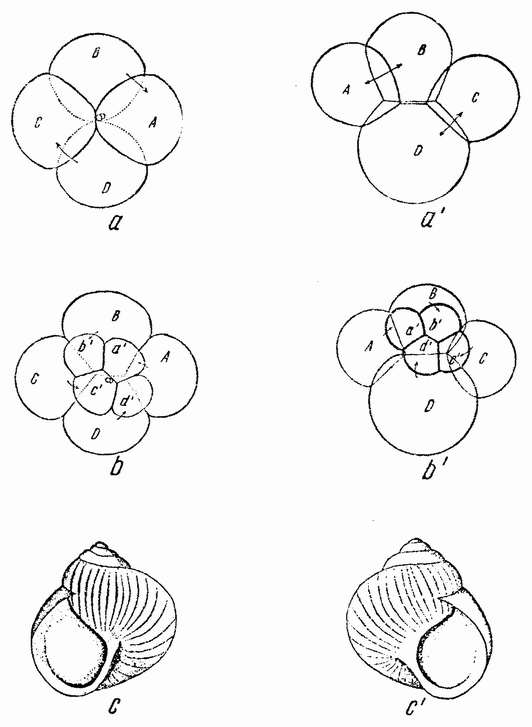 |
| Рис. 78. а, b, с — левый тип дробления и влево закрученная спиральная раковина моллюска; а1, b1, с1 — прапый тип дробления и вправо закрученная спиральная раковина моллюска (по Конклину). |
стральных потомков. Так как эти улитки могут оплодотворяться также и перекрестно, то при скрещивании декстральной улитки с декстральной или синистральной с {133} синистральной декстральная самка генетического строения ll (возникшая указанным выше путем) будет иметь возможность скреститься с декстральной же, но строения LL. Все потомство такой соматически декстральной самки будет синистральным, так как до начала дробления яйцо подвергалось влиянию двух рецессивных генов (ll). Эти синистральные улитки (Ll) в свою очередь будут давать только декстральных потомков, потому что доминантный фактор L в яйце определяет тип дробления последнего. Очевидно поэтому, что декстральные особи определенного
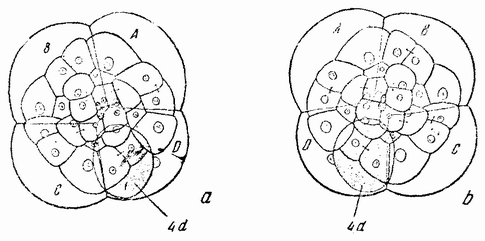 |
| Рис. 79. а — более поздняя стадия дробления яйца моллюска, в которой с правой стороны обособилась 4d клетка; b — то же самое с 4d клеткой с левой стороны. |
происхождения будут давать только синистральные семьи, а некоторые синистральные — только декстральное потомство. Наследование здесь идет согласно Менделю, но проявление признака запаздывает на одно поколение. Результат этот своеобразен потому, что симметрия взрослого организма определяется не его собственной генетической конституцией, а конституцией еще несозревшего яйца, из которого он возник. Здесь нет противоречий с обычным менделевским наследованием признаков взрослого организма, если, а это кажется правильно, симметрия определяется строением яйца перед выделением полярных телец и будучи однажды детерминированной, не может впоследствии измениться в обратном направлении независимо от того, какое генетическое строение будет иметь зигота. Что яйцо построено именно так, видно из первых стадий дробления. На четырех- и восьмиклеточной стадиях спираль дробления имеет одно направление у влево закрученных (рис. 78 a, b, c) и противоположное у вправо закрученных {134} рас (рис. 78 а1, b1, с1). Когда появляется мезодермальная клетка (4d), то она оказывается с одной стороны от проспективной средней линии у одного типа и с противоположной — у другого (рис. 79 а, b).
Все генетические данные приводят к выводу, что признаки, развивающиеся в протоплазме, в конце концов оказываются зависящими от генов в хромосомах. Более того, они показывают, что протоплазма яйца, образовавшаяся под влиянием одного набора генов, может со временем быть измененной в протоплазму, характерную для другого набора генов, привнесенного ядром сперматозоида. В случае реципрокных гибридов видно, что два набора хромосом, происходящих от двух различных рас, вызывают один и тот же конечный результат (если имеется достаточно времени для их действия) независимо от того, какая протоплазма присутствовала сначала.
С другой стороны, имеется несколько случаев, в которых реципрокные гибриды не идентичны. Кроме случаев элиминации хромосом в одних скрещиваниях и отсутствия ее в других, можно иногда ожидать различий другого рода. Так, например, случай с моллюсками показывает, что очень ранние стадии дробления по своим признакам оказываются материнскими.
Если бы эти начальные стадии влияли на все или некоторые из более поздних, то можно было бы предположить, что как своего рода последействие начальных стадий получится различный конечный результат. Если это так, то реципрокные гибриды могли бы быть различными, что и имеет место в действительности в некоторых скрещиваниях с вправо и влево закрученными улитками. Точно так же у млекопитающих, у которых плод получает питание от матери, можно было бы ожидать, что реципрокные гибриды обнаружат при рождении различия, обусловленные различиями в среде, в которой они развивались.
В связи с этим остается обсудить довольно существенный вопрос. Можно ожидать,что только в тех случаях оказывается возможным получение реципрокных гибридов, достигающих взрослой стадии, в которых протоплазма обоих родителей в. основном подобна, поэтому рискованно переносить доказательства, вытекающие из этих фактов, на все остальные случаи. Ибо если имеются реальные различия в протоплазме различных видов, не зависящие от ядерных влияний в том смысле, что они не могут быть обращены {135} одно в другое влиянием генов, то нельзя ожидать развития организма до взрослого состояния или до последних эмбриональных стадий. Поэтому до тех пор, пока не получены более обширные данные, затронутая центральная проблема не может считаться разрешенной для всех случаев.
Вопрос об относительной роли генов и протоплазмы, иллюстрированный вышеприведенными примерами, соприкасается с другим вопросом, дискутировавшимся в прошлом, а именно, что протоплазма определяет основные признаки особи, а гены только более поверхностные из них. Необходимо лишь напомнить, что опыты с влево и вправо закрученными улитками показали, что гены могут в течение времени изменить одну форму протоплазматической реакции в другую. Если мы попытаемся определить, какой из этих признаков является основным, а какой поверхностным, то наверное реверзия симметрии будет с уверенностью отнесена к первой категории признаков. Наследование четырех групп крови у человека, составляющих тройную аллеломорфу, будет также отнесено к случаям, в которых основной признак определяется генным влиянием. Наоборот, если пигмент рассматривать как поверхностный признак, то случай с шелковичным червем показывает, что он тоже может определяться протоплазмой яйца, но в последующих поколениях может быть изменен влиянием генов.
СТАБИЛЬНОСТЬ ДИФЕРЕНЦИРОВАННЫХ ТКАНЕЙ
В течение долгого времени было известно, что при некоторых условиях уже диференцировавшиеся клетки, выполнявшие определенные функции во взрослом индивидууме, могут полностью или частично передиференцироваться и затем играть другую роль в построении новой особи. Так, например, ткани некоторых асцидий в определенное время года могут образовывать шарообразные вздутия, состоящие из вещества, в котором исчезли все признаки взрослой диференцировки и из которого впоследствии может развиться новая особь. Здесь нет ясной непрерывности между типом клеток старых и новых органов. Ткань губок и гидроидов может быть протерта через мельничный газ и если затем она будет помещена на дно сосуда с морской ведой, то в результате соединения клеток, разъединенных протиранием, может образоваться {136} несколько новых особей. В какой степени любая клетка может быть частью нового организма или в какой мере ее первое состояние может влиять на ее последующую судьбу, не вполне ясно, но тем не менее в некоторых клетках должна произойти большая передиференцировка.
Некоторые опыты с прививками частично дают обратную картину. Если кусок кожи со спины только что вылупившегося цыпленка, принадлежащего к породе, отличающейся особенностями оперения, трансплантиру-
 |
| Рис. 80. Недавно привитые цыплята (по Депфорду). |
ется на то же место цыпленку другой породы (рис. 80), кожа которого в этом месте предварительно удалена, то новые перья привитого участка оказываются подобными перьям той породы, от которой происходит привитой кусочек (рис. 81). Во время операции у цыпленка имеется только пух, но фолликулы перьев уже заложены и, как установлено выше, на новом хозяине они дают начало тому же самому типу перьев, который бы они выработали, если бы остались на своем месте. Более того, если они будут выдернуты или если они вылиняют, то развивающиеся новые перья происходят из новых фолликул, образованных из старых, и сохраняют свой тип. Здесь генетическое строение отдельных клеток определяет признаки оперения привитого участка кожи, даже когда он оказывается лежащим во внешней среде хозяина, имеющего {137} совершенно отличные перья. Передиференцировки в этих условиях не происходит.
С другой стороны, было показано, что некоторые черты оперения подвержены влиянию внешних условий или специфичному воздействию внутренней секреции хозяина.
 |
| Рис. 81. Взрослые куры с прививками на крестце, а — красная курица с прививкой от петуха попосатого плимутрока; b — петух род айленд с прививкой от петуха полосатого плимутрока; с — петух рыжий лехгорн с прививкой от джерсейской белой гигантской курицы; d — курица, белый лехгорн с прививкой от курицы красный род айленд (по Денфорду). |
Так, например, если кожу с крестца молодого самца плимутрока привить на красную самку, то перья оказываются подобными соответствующим перьям самки плимутрока, а не самца (рис. 81 а). Другими словами, внутренняя секреция самки определяет некоторые признаки нового оперения. Крестцовые перья у петухов длинные, узкие и {138} заостренные, у самок же они в этом месте короткие и закругленные.
Обратная прививка обнаруживает сходный эффект. Если, например, кусок кожи с крестца черной курицы привит на спину красно-желтого петуха лехгорна, то новое черное оперение будет подобным длинным заостренным перьям петуха (рис. 81 с). Здесь этот результат объясняется отсутствием внутренней секреции, имевшейся у самки, что видно из того, что при удалении у самки яичника, ее новые крестцовые перья становятся подобными перьям самца.
Эти результаты согласуются с точкой зрения, что в случаях, когда протоплазма однажды уже была диференцирована, конечный продукт этим определен даже тогда, когда новые клетки образуются из исходных клеток, так сказать, уже направленных по определенному пути на ранней эмбриональной стадии. Результаты, касающиеся признаков окраски оперения, показывают, казалось бы, что гены не были изменены. С другой стороны, поведение в этих опытах признаков формы перьев показывает, что внутренняя среда, т. е. гормоны, вырабатывающиеся в других частях тела, тоже действует на некоторые конечные стадии диференцировки. Гормоны могут действовать сначала на протоплазму фоликулярных клеток перьев и далее посредством вызванного таким образом эффекта на продукт гена, или же они могут действовать непосредственно на гены. В настоящее время нет путей для определения, какое из этих воздействий имеет место в действительности. Имеющиеся данные не являются также решающими для вопроса, действуют ли все гены одинаково или одни из них активнее других.
Сравнение эмбрионального развития цветных пятен у пегих животных с наследованием этого же самого цвета, свойственного другим расам с одноцветной окраской шерсти, может быть обманчивым. Существуют морские свинки серые (агути), черные, красные и белые, причем наследование этих окрасок объясняется различиями в единственном гене. Имеются также пегие морские свинки. Некоторые из них могут иметь одновременно серые и белые участки шерстного покрова, другие — красные и белые, третьи — черные и белые. На первый взгляд может показаться парадоксальным, что морская свинка, у которой могут развиваться черные участки, будет иметь белые пятна в случае, если клетки обоих участков будут содержать все гены. В {139} тех случаях, где пятна точно локализованы, т. е. распространяются на определенные области тела, их локализация может не казаться отличной от какой-либо другой соматической локализации; но когда, как это имеет место у некоторых пегих животных, черные и белые пятна распределяются случайно, отмеченный выше парадокс более очевиден.
Генетические данные показали, что у мышей, крыс и морских свинок имеются определенные гены пегости. Когда присутствуют два таких рецессивных фактора, животное оказывается пегим, т. е. оно имеет участки белого и какого-либо другого цвета; например, серого. Согласно нашему предположению, гены серого цвета присутствуют во всех клетках; почему же тогда серый цвет проявляется не везде? Этот вопрос требует рассмотрения.
Для образования пигмента должно присутствовать по крайней мере два вещества; если одно из них отсутствует в клетке или не вырабатывается в ней, то окраска не развивается. В присутствии генов пегости ген серого цвета в белых участках теряет способность вырабатывать необходимый для этого цвета энзим или же протоплазма клеток некоторых участков тела не соответствует продукту таких генов. И та, другая альтернатива приводит нас к, повидимому, неразрешимой дилемме, ибо при первом предположении нельзя объяснить что подавляет ген, а при втором, повидимому, необъяснимо, почему эти протоплазматические различия находятся не на всем теле.
Эти трудности объяснения не встречаются по отношению к чисто белым но тем не менее содержащим гены серой, красной или черной окраски животных. Здесь предполагается, что в присутствии нормального аллеломорфа рецессивного фактора белого цвета вырабатывается одна из субстанций, необходимых для развития окраски. Рецессивный фактор этого свойства лишен, и развития окраски поэтому не происходит.
Здесь надо подчеркнуть, что фактор пятнистости вызывает свой эффект не тем же путем, что и фактор одноцветной белой окраски, поэтому незаконно постулировать, что взаимоотношения энзимов в обоих случаях одинаковы; в действительности они должны быть различными. Следовательно, вопрос о том, каким образом фактор пятнистости вызывает свой локализованный эффект, остается еще открытым. {140}
Существуют некоторые опыты по прививкам участков кожи у морских свинок, показывающие, что если эффект однажды был вызван, то результат не может быть изменен в обратном направлении путем перемены местоположения белого и черного участков. Когда участок кожи с черной шерстью переносится на белый участок той же самой свинки, шерсть остается черной; наоборот, белый участок на черном тоже остается белым. Тот же результат получается с черным и серым, серым и красным и т.д. Очевидно, здесь уже не локализация пятна, все равно белого или черного, определяет его цвет, ибо, будучи однажды определенным, он оказывается зафиксированным навсегда.
Это еще раз ставит вопрос, действительно ли диференциация черных и белых участков была определена их локализацией в эмбрионе или какими-либо диференциаторами, вроде, например, саморазмножающихся протоплазматических элементов, неправильно распределившихся во время раннего развития. Это приводит к положению, вновь возвращающему нас к проблеме локализации. Например, у всех серых морских свинок шерсть на спине совершенно отлична как по цвету, так и подлине от шерсти на брюхе. Если участок кожи с брюха пересажен на спину, он сохраняет все признаки брюшной шерсти. Если, как это обычно предполагают, различия между спиной и брюхом являются проблемой эмбриональной локализации, то почему мы должны воздерживаться от пользования этой же аргументацией для пегих животных, хотя бы даже, вследствие неправильности в распределении пятен, проблема казалась бы несколько более трудной?
| << | | {141} | | >> |