Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА X | ||||||||||||||||||||||||||||
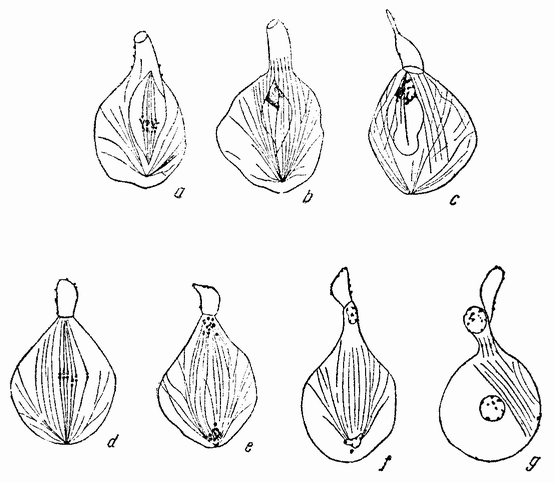 |
| Рис. 72. Первое (а—с) и второе (d—g) деления сперматоцитов у пчелы (по Мевесу). |
клеток у самцов пчелы, являющихся гаплоидами. У взрослого самца величина клеток оказалась для каждого органа соответственно одинаковой с таковой у диплоидной самки, однако имеются некоторые данные, хотя и недостаточные для того, чтобы быть уверенным в них, показывающие, что на ранних стадиях развития самца число хромосом у него может удвоиться. Если в клетках тела самца хромосомы каким-то образом удваиваются, то в его половых клетках этого не происходит, что видно во время делений {118} созревания, во время которых присутствует резко выраженное гаплоидное число хромосом (рис. 72).
У мхов и папоротников, и в меньшей степени у семенных растений, имеется гаплоидное, или гаметофитное поколение, и диплоидное, илиспорофитное, которые чередуются в жизненном цикле (рис. 73). Обе стадии настолько различны,
 |
| Рис. 73. Жизненный цикл мха. Мицелиальная нить и растение мха образуют гаплоидное или гаметофитное поколение; стебелек и капсула (с содержащимися в ней спорами) возникающими после оплодотворения, представляют собой диплоидное, или спорофитное поколение (по Маршалю). |
| | {119} | | ||
| | | |||
| | | |||
 |
| Рис. 74. Схема, показывающая образование диплоидных (2n) особей, путем регенерации спорофита у гермафродитного вида (по Маршалю). |
Последние примеры ясно показывают, что при наличии удвоенного или учетверенного набора хромосом соответственно увеличено общее количество протоплазмы. Опыты показывают также, что переход от спорофита к гаметофиту обусловливается не присутствием одного или двух наборов хромосом.
Второй вопрос касается равновесия генов. Это выражение предполагает, что общая активность генов как раз такова, что она создает характерные черты особи. Это положение может казаться тривиальностью, но истинное
 |
| Рис. 75. Хромосомы самки и самца Drosophila melanogaster (по Добржанскому). |
значение его становится ясным, когда оно рассматривается в связи с изменениями в числе генов. Характерны два рода изменений: во-первых, все диплоидное число хромосом может быть либо умножено, либо уменьшено вдвое. Равновесие, следовательно, остается неизменным, и в этом случае можно ожидать нормального результата. Но если к полному набору хромосом добавляется одна, две или больше лишних, то равновесие изменяется. Некоторых генов становится относительно больше, чем было раньше. Примеров такого нарушения равновесия известно очень много. Иногда, например, при делении половой (или любой другой) клетки, половинки какой-нибудь хромосомы могут не разойтись к противоположным полюсам, в результате чего одна дочерняя клетка получает одну лишнюю хромосому, а в другой этой хромосомы недостает. Случись это, как это и бывает иногда, в половой клетке, то в {121} оплодотворенное яйцо будет привнесена нехватка, или дупликация хромосомы, и возникает особь, являющаяся «неуравновешенной». Такая особь может выжить и тогда она обычно обнаруживает заметные отличия от исходного типа. С другой стороны, такая особь, имеет ли она на одну хромосому больше или меньше, может и погибнуть, потому что неуравновешенное отношение генов приводит к неправильностям в ее развитии. Например, Drosophila melanogaster имеет четыре пары хромосом. Отсутствие одной из самых маленьких хромосом или прибавление третьей такой хромосомы влияет на особь так слабо, что она во всем напоминает нормальную муху, за исключением лишь немногих мелких различий. Но отсутствие одной из больших хромосом или прибавление третьей такой хромосомы влияет на развитие так сильно, что эмбрион погибает.
Явное исключение имеет место в случае половых хромосом. У некоторых форм самки имеют две Х-хромосомы, а самцы только одну и, несмотря на это, самцы и самки почти по всем их признакам очень похожи друг на друга. У дрозофилы самки имеют две Х-хромосомы, а самцы одну Х-хромосому и, кроме того, вторую половую хромосому, называемую Y-хромосомой. Было показано, что в Y-хромосоме отсутствуют почти все гены, присутствующие в X-хромосоме. Можно сказать поэтому, что в таких случаях должно было установиться некоторого рода компенсирующее равновесие, заключающееся в том, что и одна, и две Х-хромосомы дают одинаковый результат, за исключением, конечно, лишь того, что две Х-хромосомы образуют самку, а одна — самца; это различие обусловлено различием в равновесии между Х-хромосомами и остальными хромосомами.
Следующие примеры иллюстрируют зависимость пола от равновесия генов. Тетраплоидная самка дрозофилы имеет формулу 4A + 4X, а нормальная диплоидная самка — 2А + 2X. Буква А в этих формулах служит для обозначения всех остальных хромосом, кроме X. Равновесие одинаково у обоих. Тетраплоидная самка при скрещивании с нормальным самцом (2A + X) дает триплоидных самок (3A + 3X), у которых равновесие тоже такое же, как у нормальной самки. В созревающих половых клетках этой триплоиднои самки хромосомы распределяются неправильно и она поэтому полустерильна; но среди немногочисленного потомства, получающегося при скрещивании ее с {122} нормальным самцом, возникает небольшое число триплоидов, несколько больше диплоидов и немного интерсексов. Последние имеют три набора аутосом (ААА) и две Х-хромосомы (3А + 2Х). Хотя такой интерсекс имеет то же число Х-хромосом, что и нормальная самка, но других хромосом он имеет больше. Это нарушение равновесия приводит к возникновению особи в некоторой степени промежуточной между нормальными самцом и самкой. Если мы предположим, что Х-хромосома содержит больше генов, определяющих женское состояние, а другие хромосомы содержат больше генов мужских признаков, то у интерсекса равновесие оказывается промежуточным. Протоплазма яйца, являющаяся во всех этих случаях одинаковой, не играет существенной роли в этом результате.
У дурмана (Datura) оказалось возможным получить целый ряд типов, в каждом из которых имеется третья хромосома той или другой пары. Добавочная хромосома приводит к образованию у особи заметных отличий как от исходного типа, так и от каждой из других форм. Могут быть прибавлены также и две хромосомы сверх нормы, что опять-таки приводит к образованию определенного изменения.
Иногда ставится вопрос, является ли нарушение равновесия при прибавлении одной лишней хромосомы менее вредным у видов с большим числом хромосом, чем в случаях, когда хромосом меньше. Такого различия, вообще говоря, можно ожидать, но прежде чем какое-либо подобное обобщение сможет иметь большое значение, необходимо учесть индивидуальные особенности видов.
Третий вопрос касается поведения хромосом, когда они на стадиях созревания конъюгируют, т. е. соединяются в пары. При таком удвоении числа хромосом, какое имеет место у тетраплоидов, имеется по четыре хромосомы каждого сорта. Обычно они соединяются в пары, которые расходятся и делятся так же, как и при правильном созревании. Каждая из четырех получающихся клеток содержит двойное число хромосом и после оплодотворения тетраплоид повторяет себя. Такой тетраплоид является саморазмножающимся. Известно несколько случаев, в которых путем скрещивания двух видов получен гибридный тетраплоид, с одним набором хромосом одного вида и одним другого. Например, были скрещены два вида Primula, у которых каждая половая клетка имеет девять {123} (гаплоидное число) хромосом; гибрид имел восемнадцать хромосом. Это было мощное растение, но при созревании его половых клеток хромосомы, происходящие от двух видов, не могли правильно соединяться в пары, и поэтому зрелые половые клетки оказывались содержащими различные числа хромосом и были неполноценными. Этот гибрид был поэтому стерилен. Но у него появилась ветка с удвоенным числом хромосом. Несомненно, что эта ветка возникла благодаря тому, что протоплазма одной клетки не разделилась после разделения хромосом, в результате чего получилась тетраплоидная клетка, имеющая по две хромосомы каждого сорта от обоих видов. Из этой клетки и возникла тетраплоидная ветка. На этой ветке образовались цветы, которые были фертильными, потому что хромосомы могли теперь соединяться в пары, ибо хромосомы одного вида соединялись с им подобными, а хромосомы другого вида тоже имели себе партнеров. Потомство оказалось тетраплоидным гибридом, и полученный новый вид плодовитым.
Если тетраплоид скрещен с диплоидом, то потомство будет триплоидным и, как правило, имеет не полноценные половые клетки. Потомство это стерильно благодаря тому, что все три сходные хромосомы не могут соединиться в пары, что при созревании половых клеток приводит к неправильностям в распределении хромосом. Случается, однако, что половая клетка может получить гаплоидный набор хромосом. Если соединяются две такие клетки, то диплоидное состояние опять восстанавливается. Другие возможные комбинации дают неуравновешенные типы потомков.
Как было описано выше, тетраплоидные клетки могут появляться у растения путем неразделения протоплазмы при расхождении хромосом. И действительно, в растительных тканях иногда встречаются тетраплоидные клетки. Такие клетки могут быть получены искусственно, путем воздействия на клетку во время процесса деления, агентом, препятствующим следующему клеточному делению. Дочерние хромосомы образуют тогда одно ядро с удвоенным числом хромосом, и это число сохраняется в течение всех последующих делений. Изредка могут также возникнуть и половые клетки с диплоидным числом хромосом вместо гаплоидного. Не может быть сомнений, что такие клетки получаются в результате подавления деления протоплазмы. {124} Такие тетраплоидные половые клетки крупнее нормальных.
Естественно, напрашивается вопрос, останется ли клетка крупнее после дальнейших делений, если к ней искусственно прибавить протоплазмы; и если это так, то останутся ли теми же размеры хромосом. В действительности было показано, что на некоторых стадиях дробления, после того как клетка разделилась на одну большую и другую меньшую, хромосомы в последней мельче, чем в первой. Являются ли гены затронутыми таким же образом, неизвестно. В настоящее время на этот вопрос не может быть дано ответа, но если бы общее количество протоплазмы могло быть искусственно увеличено, то это служило бы доказательством наличия в ней саморазмножающихся единиц, не зависимых от генов. Так как известно, что гены влияют на общее количество протоплазмы, как, например, это имеет место у тетраплоидов, то ясно, что между генами и протоплазмой существует некоторое взаимоотношение и, нужно думать, что предложенный выше опыт в случае отрицательного результата не пролил бы никакого света на этот вопрос.
Имеются некоторые данные об обратном положении вещей, когда гаплоидные особи или гаплоидные клетки появляются у диплоидной особи. В этом случае у гаплоидных растений отдельные клетки, повидимому, мельче, чем у диплоидов. У гаплоидных самцов животных, подобных коловраткам, тлям и пчелам, имеющиеся в настоящее время данные недоказательны. Гаплоидные участки случайно были найдены у диплоидных особей дрозофилы. Клетки в таких участках мельче. Является вероятным, что в других случаях общее количество протоплазмы коррелировано с числом хромосом и может показаться, что количество протоплазмы зависит от генов, но вопрос о том, имеются ли в протоплазме саморазмножающиеся единицы, которые наследуются независимо от единиц в хромосомах, или же увеличение, или уменьшение количества протоплазмы зависит от генных отношений, все же остается открытым.
| << | | {125} | | >> |