Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА VI |
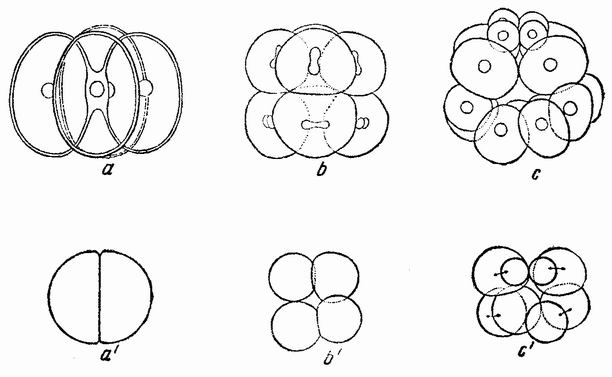 |
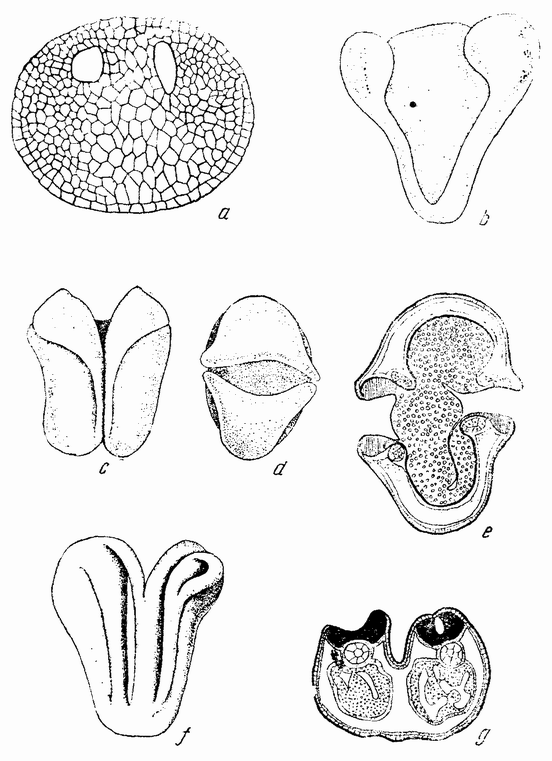
| Рис. 40. а, b, с — четырех-, восьми- и шестнадцатиклеточная стадии яйца морского ежа; а1, b1, с1 — первые три дробления изолированной половины бластомера (по Дришу). |
делился бы, если бы он оставался соединенным со своим соседом, т.е. борозда проходит через анимальный полюс (рис.40, a1). Следующее деление является экваториальньш, а следующее за ним образует у вегетативного полюса микромеры (рис.40, b1—с1). При первых делениях открытая сторона {75} половинной бластулы закрывается и, по мере того как продолжаются деления, внутри бластулы появляется пространство, заполненное жидкостью. Затем происходит га-
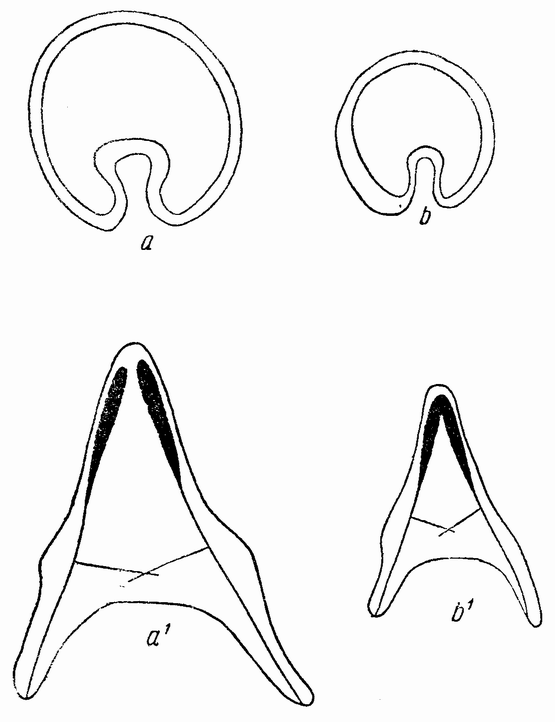 |
| Рис. 41. Целые и половинные гаструлы и плутеусы морского ежа; последние развились из половинных бластомеров. |
струляция, после чего образуется типичный плутеус (рис. 41, b—b1).
Способ закрытия бластулы и инвагинации первичного кишечника представляет в некоторых отношениях {76} теоретический интерес. Эти процессы могут быть изучены путем окраски вегетативной области половины шестнадцатиклеточной стадии. Полученные результаты показывают, что открытая сторона половины бластулы закрывается и клетки анимального и вегетативного полюсов (равно как и клетки с боков бластулы) сближаются. Инвагинированный первичный кишечник начинается в середине энтодермального поля. Эти отношения показаны на рис. 42, на котором изображена целая бластула (b), ее нормальный способ
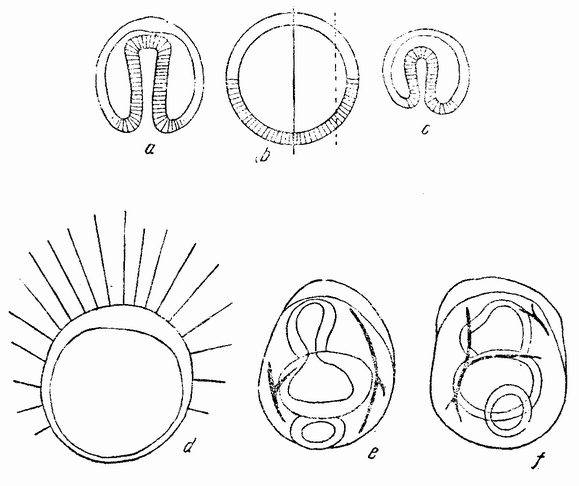 |
| Рис. 42. b — нормальная бластула и способ ее инвагинации (а), с — для сравнения инвагинация половинной бластулы (по Герстадиусу). |
гаструляции (а) и гаструляция половинной гаструлы (с). Инвагинация происходит симметрично, причем впячивается только энтодермальное вещество. Это, казалось бы, означает, что первоначальный центр энтодермального вещества (которое после разделения лежит на стороне половинной бластулы) не является центром гаструляции половинной бластулы, а что этот центр восстанавливается в середине энтодермального вещества. Это смещение центра можно представить следующим образом. Клетки на анимальном полюсе приходят в соприкосновение с клетками {77} вегетативного полюса, где сначала был центр энтодермы. Если затем, благодаря этому соприкосновению, энтодермальные клетки теряют некоторые из своих способных к диффузии веществ, и, наоборот, эктодермальные клетки отдают энтодерме некоторые из своих веществ, то наиболее концентрированное вещество энтодермы окажется лежащим ближе к середине энтодермальной массы. Образуется новый центр, который становится центром гаструляции. Если на четырехклеточной стадии яйца морского ежа изолировано четыре бластомера, то каждый сегмент является частью целого. При втором делении образуется только единственный микромер. Развивается бластула вчетверо меньшей величины, которая гаструлируется и превращается в эмбрион-плутеус, величина которого тоже вчетверо меньше. Правда, не было обнаружено, что все четыре бластомера одного и того же яйца ведут себя одинаковым образом, но надо думать, что это так.
Надо отметить, что как половинки, так и четвертушки бластомеров содержат все вещества яйца от анимального до вегетативного полюсов, но после третьего деления четыре верхние клетки оказываются содержащими только анимальное вещество, а нижние клетки только вегетативное. В действительности было обнаружено, что некоторые из изолированных восьмушек бластомеров гаструлируют, тогда как другие остаются плавающими бластулами. Однако потенции верхних и нижних клеток лучше всего видны при отделении четырех верхних бластомеров от четырех нижних. Было найдено, что четыре верхние (анимальные) клетки дают начало плавающей бластуле (рис. 42, d), которая не гаструлирует, но у которой может произойти инвагинация рта. Четыре нижних клетки дают начало гаструле, у которой впоследствии развивается пищеварительный тракт, разделенный на характерные три отдела. Может происходить и начало образования скелета, но рот все же не образуется (рис. 42, е—f). Из этого очевидно, что верхняя и нижняя половины развиваются только в частичные структуры, сходные с теми частями, которые развиваются из этих же самых участков целой бластулы.
ОПЫТЫ С ЯЙЦАМИ АСЦИДИИ
У асцидии положение вещей несколько отлично от такового у морского ежа, ибо, хотя у обоих дробление резко неравномерное, у асцидии гаструляция начинается на {78} более ранней стадии, причем она сначала приближается к типу, дающему половинную гаструлу, и лишь затем этот процесс до некоторой степени выравнивается.
С помощью встряхивания сегментирующихся яиц можно убить один из бластомеров, не повреждая других. В этом случае поврежденный бластомер действительно убит, а не просто устранен от развития, как это имело место в опытах с яйцами лягушки, хотя в некоторых яйцах это
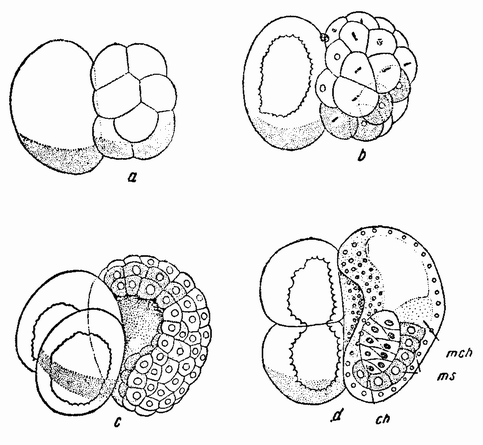 |
| Рис. 43. Дробление, гаструляция и образование зародыша из половинного бластомера Styeia. Поврежденный бластомер не мертв, но его развитие запаздывает. Он еще соединен с более развитой половиной (по Конклину). |
все же может случиться. В одном опыте бластомеры были разделены полностью, и было показано, что они ведут себя таким же образом, как в тех случаях, когда один из них убит. Клетка за клеткой, изолированная половина бластомера претерпевает те же самые деления, которые претерпела бы эта половинка, если бы она осталась соединенной с другим живым бластомером (рис. 43). Поэтому в то время, когда зародыш начинает гаструлировать, половинка содержит как раз половину того числа клеток, которое должно быть в целом эмбрионе, и гаструляция {79} происходит, если можно так выразиться, только с одной стороны вокруг рта гаструлы, а не со всех сторон его, как это имеет место в целом эмбрионе. В тот момент, как только инвагинировались клетки первичного кишечника, спинной струны, мускулатуры и начинают сближаться поверхностные клетки, предназначенные для образования нейральной пластинки, наблюдается замечательный факт. Первичный кишечник оказывается не половинной, а цельной округлой полостью, спинная струна образует единый закругленный валик клеток, а нейральная пластинка образует единую билатерально-симметричную структуру. Другими словами, некоторые органы оказываются не половинными структурами, а вполне или почти целыми. С другой стороны, клетки, предназначенные для образования мускулатуры, не дают правых и левых структур, а образуют их только с одной или с другой стороны зародыша в зависимости от того, произошел ли эмбрион из правого или левого бластомера. Неспособность предетерминированных мускульных клеток вести себя так же, как ведут себя и другие клетки, а именно образовывать целый орган половинной величины, может быть правдоподобно объяснена их положением во время инвагинации клеток спинной струны. Мускульные клетки лежат с одной стороны спинной струны и для них может быть механически затрудненным передвигаться вокруг спинной струны для образования мускулатуры с другой стороны ее. Как бы то ни было, факт остается фактом, что дробления, подобные дроблению яиц морского ежа, находятся в строгом соответствии с той частью яйца, в которой они происходят, и эмбрион, развившийся из любой из первых двух клеток, значительно более похож на целую структуру, чем на половинную. Во многих отношениях он развивается совсем таким же образом, как и зародыш морского ежа. В обоих случаях развитие связано с сильно выраженной саморегуляцией.
Если у асцидии на четырехклеточной стадии изолированы две клетки, лежащие на дорзальной стороне яйца, на которой располагается полумесяц, то эти клетки продолжают сегментироваться, как будто бы они являются частью целого (рис. 44, а, b). Эмбрион гаструлируется, но после закрытия бластопора его развитие заканчивается. Нейральная пластинка присутствует в полной мере, но не заворачивается для образования трубки, Клетки спинной {80} струны тоже присутствуют, но не удлиняются для образования валика. Имеется и первичный кишечник. Статическое состояние этих органов может объясняться их неспособностью вытягиваться. Эмбрион не имеет мускульных клеток, каудальной энтодермы и мезенхимных клеток.
При изолировании двух других бластомеров, лежащих напротив полумесяца, они дробятся как часть целого
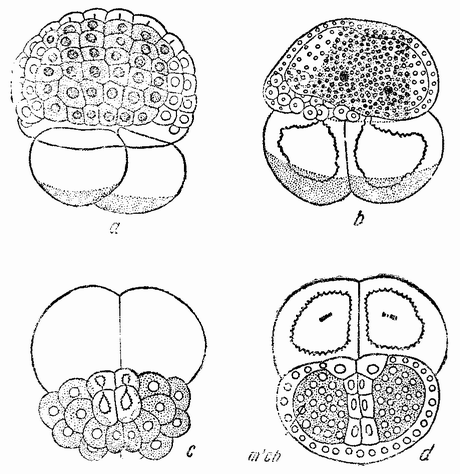 |
| Рис. 44. а и b — развитие двух передних бластомеров Styela; с и d — развитие двух задних бластомеров; m, ch — мезенхима (по Конклину). |
(рис. 44, с—d). Мускульные клетки образуются позднее. Появляется также каудальная энтодерма и мезенхимные клетки. Следы спинной струны или нейральной пластинки отсутствуют и не происходит образования хвоста. Об этих двух эмбрионах можно сказать, что один из них потому не развивается в целый зародыш или хотя бы даже в более или менее нормальную форму, что в отсутствии другой половины он не может вытянуться, а другой потому, что он не получил в придачу от второй половины спинную струну и нейральную пластинку, как это происходит с этим участком в нормальном зародыше. {81}
Дробление одной четвертушки бластомера зависит от того, из какой части квадранта она происходит, являясь в любом случае строго неравномерным. Дальнейшее развитие ее подобно таковому у только что описанных двух соединенных бластомеров.
ПОЛОВИННОЕ И ЦЕЛЬНОЕ РАЗВИТИЕ У АМФИБИЙ
В свете отчетливых результатов, полученных на яйцах асцидий, могут быть лучше поняты результаты, полученные Ру на яйцах лягушек. Более того, опыты с яйцами другой амфибии — тритона, дают дальнейшие факты, касающиеся этой же самой проблемы. Плоскость первого
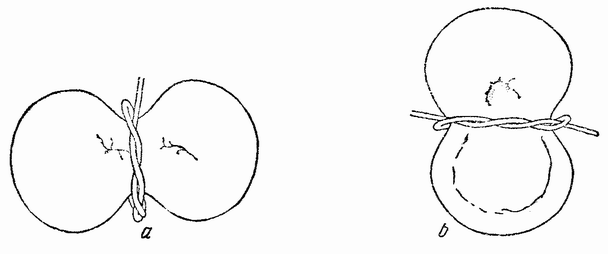 |
| Рис. 45. а — стадия гаструлы яйца тритона, которое на двухклеточной стадии было перетянуто волоском по линии будущей средней плоскости зародыша; b — подобное же яйцо, которое тоже было перетянуто на стадии двух клеток, но у которого первая плоскость дробления была фронтальной, т. е. проходила под прямым углом к средней плоскости, являющейся в яйце тритона более обычным положением плоскости первого дробления (по Шпеману). |
дробления яйца тритона обычно параллельна плоскости серого полумесяца, но средняя плоскость эмбриона соответствует не этой плоскости дробления, а плоскости борозды второго дробления, которая проходит через середину полумесяца. В некоторых яйцах тритона плоскость борозды первого дробления проходит через середину полумесяца и тогда она соответствует средней плоскости зародыша. Сначала надо рассмотреть последний случай. Если такое яйцо перевязать волоском в плоскости первого дробления таким образом, чтобы на яйце была перетяжка (рис. 45, а), и если волосок затягивать по мере дальнейшего деления яйца, то в результате разовьются два целых {82} эмбриона половинной величины. Бластопор появляется в каждой части в области половины полумесяца и, по мере того как его губы обрастают желток, вместо того чтобы следовать линии перетяжки между половинками, заворачивается в каждой части несколько в сторону от этой линии. Поверхностные клетки снаружи губ инвагинируются, как у нормальной целой гаструлы. Нейральная пластинка образуется над инвагинировавшейся хордальной энтодермой.
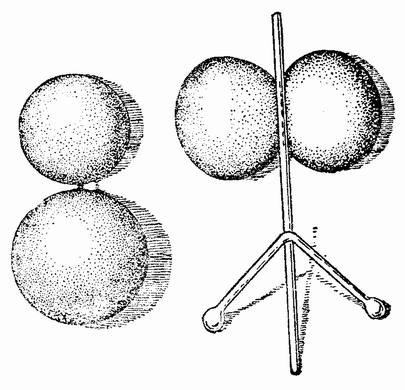 |
| Рис. 46. Стадия двух клеток яйца тритона, оболочка которого была удалена (слева). Два бластомера разделены с помощью стеклянной палочки (справа) (по Мангольду). |
Такой же точно результат получается, если вдоль плоскости первого дробления положить стеклянную палочку (рис. 46, справа), которая с помощью грузика медленно вдавливается между двух развивающихся половинок.
Иное получается, когда первое дробление происходит в плоскости, параллельной серому полумесяцу (рис. 45, b). В этом случае в половинке, содержащей полумесяц, губа гаструлы развивается в области полумесяца и образуется целый эмбрион половинной величины. Другая же половина не гаструлирует и не образует зародыша. Из этих результатов ясно, что для того, чтобы развитие шло дальше, важно наличие области серого полумесяца или хотя бы половины его. {83}
Значение этого участка показано также другими опытами, в которых сращивались вместе половинки двух яиц, каждая из которых содержит кусочек участка губы гаструлы. Например: если два эмбриона, каждый из которых находится на стадии гаструлы, разрезаны на две части вдоль средней оси и правая часть одной половины соединена с другой половиной в соответствующем же положении (т. е. сторона к стороне), то может возникнуть единый эмбрион. Но если половинки содержат целую дорзальную губу и оси обоих соединенных половинок повернуты немного в стороны друг от друга, как показано на рис. 47, а,
 |
| Рис. 47. а — соединение двух половинок гаструлы с дорзальными губами, повернутыми под углом друг от друга; b — зародыш с двумя головами и одним туловищем, образовавшийся в результате такого соединения (по Шпеману). |
то образуется эмбрион с двумя головами и одним туловищем (рис. 47, b).
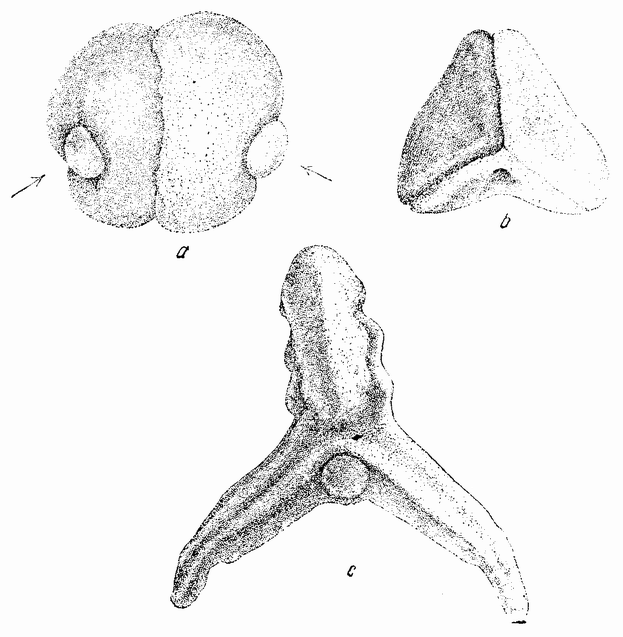
Если у обоих гаструл отрезать верхние половинки, а нижние половинки, каждая из которых содержит целую губу гаструлы, соединить вместе так, как показано на рис. 48, а, т. е. таким образом, чтобы их оси образовывали друг с другом небольшой угол (как показано стрелками), то разовьется зародыш, имеющий одну голову и два хвоста (рис. 48, b, с). Из этого следует, что вещества, инвагинированные в месте образования губ гаструлы, продвинулись вперед и слились, образуя единую структуру, тогда как на заднем конце они остались раздельными и из каждого развилась отдельная структура.
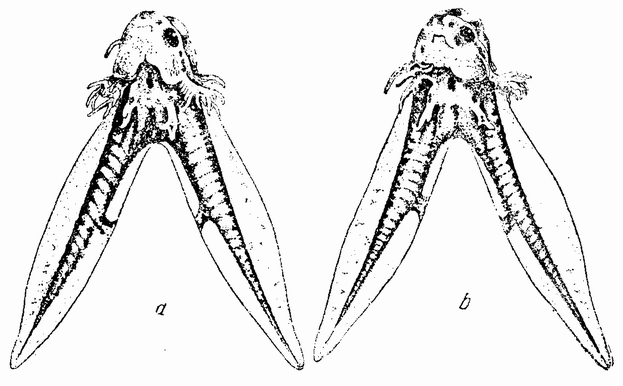
Еще более интересная комбинация получается при соединении двух половинок гаструл, у которых передние концы каждой дорзальной губы лежат как раз друг против друга {84} (рис.49, с). В этом случае образуется эмбрион с двумя головами (рис. 49, b), каждая из которых состоит из частей каждой половины, и с двумя туловищами — по одному из каждой половинки (рис. 50, а, b). Здесь инвагинирован-
 |
| Рис. 48. a — соединение двух половинок гаструлы с дорзальными губами, повернутыми под углом друг к другу, как показано стрелками; b — стадия нейрулы, развившаяся из такого зародыша; в этом случае в результате соединения материала обеих гаструл образуется единственная голова (на рисунке каждая из соединенных половинок различно окрашена); с — более взрослая стадия того же самого зародыша с одной сложной головой и двумя независимыми туловищными отделами, по одному для каждой половины (по Шпеману). |
ные материалы, двигаясь вперед, встречаются на линии соединения и одна половина каждого вывертывается наружу для образования одной головы, а другая половина — для образования другой. {85}
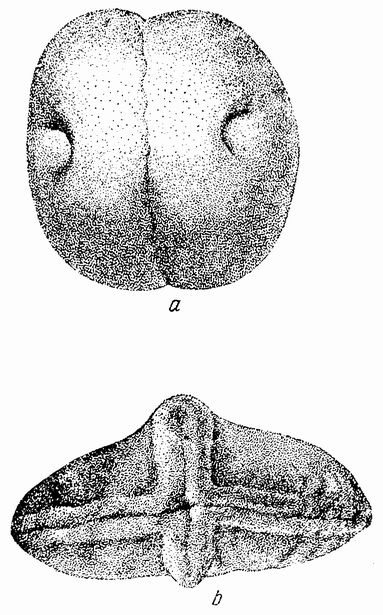
Развитие яйца лягушки настолько подобно разлитию тритона, что кажется вероятным, что если один бластомер полностью удалить, то из другого разовьется полный эмбрион. Это и происходит в действительности. В большин-
 |
| Рис. 49. а — соединение двух половинок гаструлы тритона, дорзальные губы которых направлены точно друг против друга; b — развившийся из них зародыш, имеющий две головы, каждая из которых образована из материалов обеих половинок и повернута в противоположных направлениях, и два раздельных туловищных отдела по одному из каждой половины. Срединная плоскость голов проходит под прямым углом к таковой двух туловищных участков. В результате получается типичный крестовидный зародыш (по Шпеману). |
стве яиц было невозможно отделить один бластомер, не повреждая другого, но в одном из трех опытов удалось содержимое одного из бластомеров осторожно высосать. {86}
Оставшийся бластомер развился в целый зародыш половинной величины.
Способность каждого из первых двух бластомеров яйца лягушки развиваться в целый зародыш была косвенно показана путем инвертирования яйца после наступления стадии двух клеток и удержания его в этом положении в течение последующих стадий (рис. 51, а—е). Известно, что в этих условиях вещества внутри обоих бластомеров
 |
| Рис. 50. Противоположные стороны крестовидных зародышей тритона, образованных в результате соединения двух половинок гаструл, как это изображено на рис. 49 (по Шпеману). |
сами перемещаются в соответствии с их весом. Более тяжелый желток при этом опускается вниз вдоль плоскости деления (рис. 51, b, с), отодвигая вещество серого полумесяца в одном бластомере от такового в другом. Каждая половина образует затем губу гаструлы и дает начало целому зародышу, развитию которого лишь несколько мешает соединение половинок (рис. 51, е). Описывалось несколько типов двойных эмбрионов, в зависимости от того, как раздвигалось вещество полумесяца (рис. 52). В свете этих данных можно теперь объяснить опыты Ру. Употребляемая им горячая игла не разрушала, а только повреждала вещества в той части яйца, в которую она вводилась. В этих условиях участок полумесяца остается {87} как целое и половина его инвапширует; поэтому вдоль линии соединения образуется половина эмбриона, как это происходило бы, если бы обе половинки развивались одновременно. Пожалуй, можно сказать, что участок серого полумесяца действует как цельный организатор, но только та часть его, которая превратилась в клетки, может продолжать развиваться в половинную структуру таким же образом, как это происходит в нормальном зародыше.
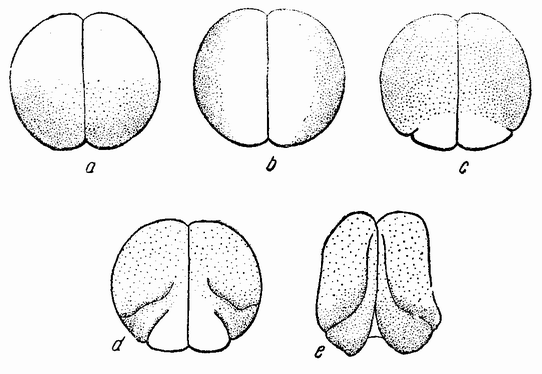 |
| Рис. 51. Схемы инвертированной двухклеточной стадии яйиа лягушки, а — яйцо после инверзии; b — погружение желтка вдоль делящейся клеточной стенки; с — дорзальная губа, появляющаяся в каждой половине; d и е — образование двух зародышей, по одному из каждой половины. Этот способ гаструляции не находится в согласии с описанным Шлейпом процессом (по Дюркену). |
ОПЫТЫ С ЯЙЦАМИ ЛАНЦЕТНИКА
После оплодотворения в яйце ланцетника образуется плоскость билатеральной симметрии. Вещество полумесяца, которое при окраске яйца окрашивается в более темный цвет, лежит на задней стороне яйца (рис. 53, ms). Этот участок яйца становится мезодермой. Желток энтсдермальных клеток лежит впереди и вокруг вегетативного полюса (рис. 53, end). Напротив полумесяца лежит вещество, из которого образуются нейральная пластинка и спинная струна. Расположение этих веществ на четырех и восьмиклеточнои стадиях показано на рис. 53, c, d. Борозда
| | {88} | |
 |
| Рис. 52. Зародыши из инвертированной двухклеточнои стадии яйца лягушки, а — разрез через стадию дробления; b — два целых зародыша с противоположных сторон яйца; с — два зародыша, соединенных спина к спине; d — то же самое с переднего конца; е — поперечный разрез через последний. У этих зародышей половинные нейральные пластинки по бокам двух эмбрионов не соединились в одну нейральную трубку, как в типичном крестообразном удвоении, и они интерпретируются как два целых зародыша со spina bifida; f — два зародыша, соединившиеся боками; g — поперечный разрез через такой зародыш (по Ветцелю). |
| | {89} | |
первого дрооления делит яйцо на две равные части; борозда второго — на две почти равные клетки и на две меньших, лежащих с задней стороны. При третьем делении четыре клетки анимального полушария оказываются мельче, чем четыре клетки вегетативного полушария (рис 53, d).
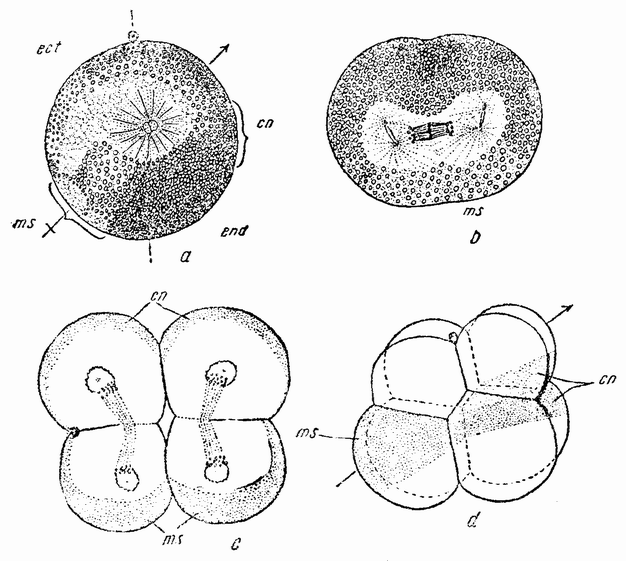 |
| Рис. 53. Разрез через среднюю плоскость яйца ланцетника. ms — мезодермальный полумесяц на заднем конце; cn — хордо-нейральный полумесяц на переднем конце; end — эндодермальная область на дорзальной стороне; ect — эктодермальный полумесяц на вентральной стороне; b — фронтальный разрез, сделанный не задолго перед первым дроблением; с — конец второго дробления, задние клетки содержат мезодермальный полумесяц, они мельче передних клеток с хордо-нейральным полумесяцем; d — восьмиклеточная стадия, четыре микромера сдвинуты вперед над четырьмя макромерами; обозначения те же, что и раньше (по Конклину). |
Изолированные бластомеры двухклеточнои стадии, как правило, дробятся как половинки и из них развивается целый зародыш половинных размеров. Изолированные бластомеры четырехклеточной стадии тоже дают начало {90} зародышу, имеющему форму целого эмбриона, но зародыш, развивающийся из бластомеров передней четверти, оказывается частично неполным в отношении одних органов, а зародыш из задней четверти — неполным в отношении других органов. Личинка, происходящая из задней части, не имеет неиральной пластинки и хорды; личинка из передней части имеет нейральную пластинку, хорду и следы сомитов, но из нее все же никогда не образуется полная личинка. Развитие изолированных бластомеров ланцетника во многих отношениях подобно таковому у асцидий.