Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА IV |
 |
| Рис. 13. Движение красного пигмента в яйце Paracentrotus во время оплодотворения (по Бовери). |
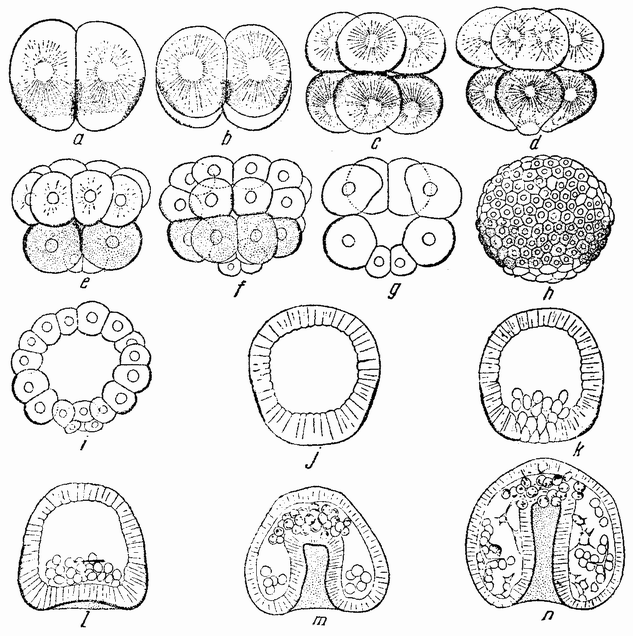
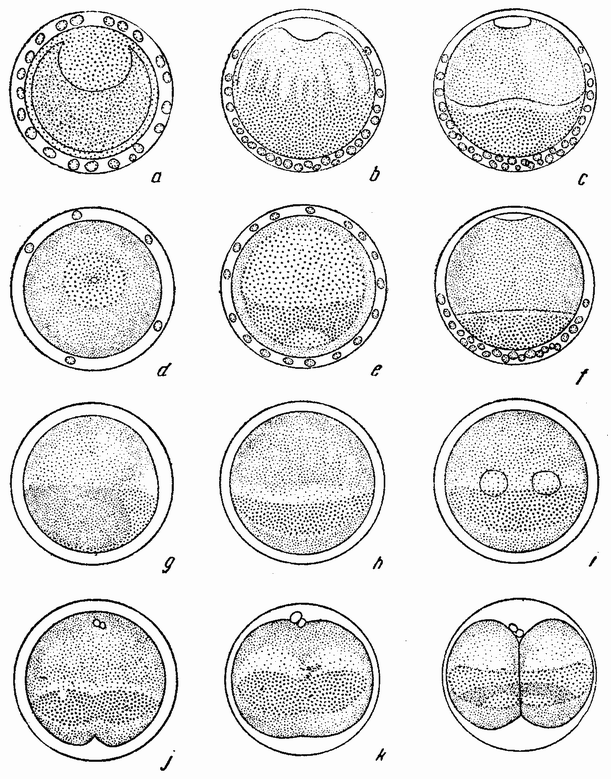
вого деления яйца морского ежа проходит через анимальный полюс (рис. 14, а), центр яйца и вегетативный полюс. Второе деление, происходящее под прямым углом к первому, тоже проходит через анимальный полюс (рис. 14, b). Третье деление проходит поблизости от экватора яйца (рис. 14, с). Восемь возникающих при этом клеток имеют приблизительно равные размеры.
Положение плоскостей дробления определяется еще до осуществления каждого деления положением веретен, которые лежат в плоскостях, перпендикулярных плоскостям будущих делений. Возникает вопрос, определяется ли плоскость деления положением веретена, где бы оно ни находилось, или же эта плоскость является фиксированной, а веретено перед делением передвигается в соответствующее положение. Данные, относящиеся к этому вопросу, получены в опытах сдавливания и центрифугирования яйц. Эти данные если и не вполне убедительны, то все же в некоторой степени дают ответ на этот вопрос. {38}
Например, путем легкого сдавливания яйца между предметным и покровным стеклами можно изменить положение плоскостей дробления; первое, второе, третье и даже более поздние дробления (рис. 15, а—е) происходят перпендикулярно к сдавливающим пластинкам. Если сплющенное яйцо окажется лежащим так, что его анимальный полюс
 |
| Рис. 14. Дробление и гаструляция яйца Paracentrotus (по Бовери). |
будет против покровного стекла, то первые два деления пройдут через анимальный полюс и может случиться, что они окажутся лежащими на том же меридиане, на котором они были бы, если бы яйцо не было сдавлено. При этих условиях первые веретена будут лежать в своих обычных положениях. Но если яйцо окажется лежащим на боку, {39} т. е. анимальный полюс его будет под углом в 45° к сдавливающей пластинке, то положение оказывается иным; в этом случае деление пройдет все же через полюс и под прямым углом к пластинкам и так как оно проходит также и через экватор веретена, то веретено должно во многих случаях сдвигаться или образовываться на новом месте. Это более очевидно при втором делении сдавленного яйца, когда плоскость дробления вовсе не проходит через полюс, а лежит под прямым углом к нему, т. е. в первоначальной
 |
| Рис. 15. Яйцо морского ежа, содержавшееся живым при продолжительном сжатии в струе морской воды (по Циглеру). |
экваториальной плоскости яйца, в которой проходит плоскость третьего нормального дробления. Из этого следует, что при сдавливании веретена смещаются и располагаются под прямым углом к направлению давления, т. е. параллельно стеклянным пластинкам. Следовательно, их положение, по крайней мере частично, определяет место, где пройдет борозда дробления.
Конечно, положение вещей является несколько более сложным, чем только что описанное, потому что веретено является лишь частью очень сложной митотической фигуры, занимающей почти все яйцо во время деления. Известно, что вся структура (фигура митоза) под влиянием сдавливания может сместиться и передвинуть собственно веретено {40} в новое положение, причем ответственной за этот результат является вся фигура деления, а не одно лишь веретено или прикрепленные к нему хромосомы. Как известно, фигура деления представляет собой, гель, более плотный, чем остальное вещество яйца, в котором она, так сказать, плавает. Она может быть механически сдвинута как целое и может, вероятно, также в некоторой степени быть перестроенной в своем новом положении. Наиболее убедительное доказательство этого получено при центрифугировании яиц.
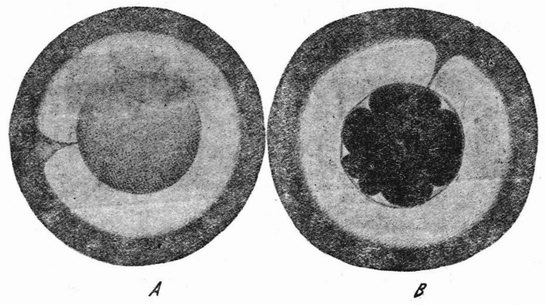 |
| Рис. 16. Центрифугированные яйца морского ежа Arbacia. A — слоистое яйцо Arbacia, окруженное прозрачной слизью; с одной стороны видна воронка. В — восьмиклеточная стадия с микромерами напротив воронки (по Моргану и Спунеру). |
Если яйца морского ежа центрифугируются в пробирке с морской водой при очень большой скорости вращения, то они сразу опускаются на дно и более тяжелые вещества яиц перемещаются к наружному или центрифугальному краю, а более легкие к центральному или центрипетальному. Все вещества располагаются при этом четырьмя слоями, как это показано на рис. 16 А. На внутреннем конце оказывается шапка масла или жира, затем идет слой прозрачной протоплазмы, за ней широкий слой, заполненный зернышками желтка, и, наконец, слой красных пигментных зерен. Если имеется сегментационное ядро, то оно лежит в прозрачном слое непосредственно под жиром. При наличии фигуры деления она тоже располагается в этой прозрачной полосе и притом всегда параллельно слоистости, причем дробление (рис. 17, a) разрезает эту слоистость {41} под прямым углом. В студенистой массе, окружающей яйцо, имеется воронка, которая может быть сделана видимой, примешивая в воду тушь. Эта воронка соответствует анимальному полюсу яйца (рис. 16 A) и этим путем может быть показано, что в центрифугированном яйце плоскость борозды первого дробления не занимает постоянного положения по отношению к полюсу. Из этого следует, что положение плоскости дробления определяется местополо-
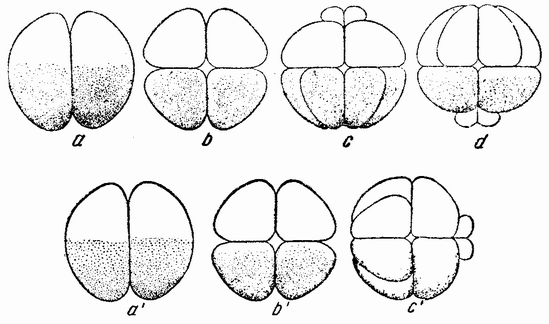 |
| Рис. 17. Дробление центрифугированных яиц Arbacia, в которых первая плоскость деления проходит под прямым углом к слоистости, а вторая под прямым углом к первой и параллельно слоистости. Третье дробление происходит в плоскости бумаги. Четвертое дробление изображено на рис. с, d и c'; микромеры могут лежать у центрипетального полюса (с), у центрифугального полюса (d) или сбоку (с') (по Моргану). |
жением веретена. Другими словами, в нормальном яйце сама фигура деления приспособляется к положению полюса и поэтому первое деление проходит через него. Плоскость деления как таковая не предопределена анимальным полюсом или во всяком случае ее положение может быть изменено. Борозда второго деления центрифугированного яйца проходит под прямым углом к первому и параллельно слоистости (рис. 17, b), что означает, что его веретено располагается под прямым углом к веретену первого дробления. Две из образовавшихся клеток будут содержать весь жир и большую часть прозрачного вещества, в других же двух клетках будет желток и пигмент. Третье деление происходит {42} под прямым углом к первым двум, образуя четыре клетки одного рода и четыре другого.
Представляет интерес следующее дробление, при котором в точке пересечения двух плоскостей деления возникают четыре микромера (рис. 17, с—d). Они могут лежать в месте пересечения плоскостей первого и второго или первого и третьего деления в пигментном или в желточном полях, или между ними. Было найдено, что где бы они ни располагались, микромеры образуются напротив воронки или возможно более близко от нее. Из этого, повидимому, следует, что ко времени появления микромеров в яйце имеется область, дающая им начало, и что образование их при четвертом дроблении в высокой степени независимо от последовательности появления предыдущих плоскостей.
На более поздней стадии микромеры перемещаются внутрь бластулы (рис. 14, k—n) и превращаются в мезенхиму. Может казаться поэтому, что в яйце имеется определенная область, которой предначертано стать микромерами; но центрифугирование яиц не дает ответа на вопрос, локализуется ли эта область на вегетативном полюсе до или во время выделения полярных телец или, может быть, она образуется где-либо в другом месте и передвигается в определенную часть яйца после того, как оно уже оплодотворено или во время первых дроблений. Имеются, однако, другие опыты, относящиеся к этим вопросам.
Если яйца морских ежей сильно встряхнуть в небольшой пробирке, то они могут быть разломаны на части, которые затем могут быть оплодотворены. Некоторые из этих частей будут содержать пронуклеус яйца, в других же он будет отсутствовать, но как те, так и другие при проникновении в них сперматозоида будут развиваться. Образование микромеров в фрагментах очень неправильно и так как фрагменты являются различными и неизвестными частями яйца, то из этих результатов не может быть сделано достоверных выводов. Более точный метод заключается в разрезании яиц на две части и в прослеживании развития каждой из них. К сожалению, в большинстве яиц морского ежа пронуклеус перед оплодотворением не занимает определенного фиксированного положения по отношению к анимальному полюсу; полярные тельца теряются и воронка не может быть видна без обработки тушью. Но у вида Lytechinus полярные тельца сохраняются, что делает возможным ориентировку разреза. Если эти яйца разрезаны в {43} вертикальной плоскости, т. е. вдоль оси яйца, то после оплодотворения развиваются оба куска, один из которых оказывается гаплоидным, а другой диплоидным. Однако оба эмбриона (плутеуса) не могут быть отличены от нормальных, кроме как по своим размерам. Когда оба фрагмента делятся, внешний вид одного из них кажется нормальным, другой же фрагмент не имеет микромеров. Это, повидимому, означает, что либо разрез прошел с одной стороны оси и вегетативный полюс находится только в одном фрагменте, либо же, что вещество микромеров было в это время в центре или вблизи центра яйца и лежит с одной стороны разреза. Последующие опыты делают трудным окончательное решение, так как исследователи пришли к заключению, что в этих яйцах отсутствует локализованное вещество, образующее микромеры, — по крайней мере такое вещество не «диференцируется» перед оплодотворением. Что такое вещество может тем не менее присутствовать в яйце даже если оно и не диффузно распространено на поверхности или сконцентрировано вблизи центра яйца, не было убедительно показано. Однако существуют другие опыты, проведенные на яйцах морского ежа Arbacia, показывающие, повидимому, что такое вещество присутствует внутри яйца еще до того, как произошло оплодотворение. В связи с этим надо напомнить, что в яйцах Arbacia вещество, из которого образуются микромеры, всплывает изнутри яйца к вегетативному полюсу, когда веретена микромеров касаются этого участка. В действительности микромеры состоят почти полностью из вещества внешней полярной звезды и только тонкий поверхностный слой состоит из исходного анимального вещества.
ЯЙЦО АСЦИДИИ
Со времени первых наблюдений Эдварда Ван Бенедена над яйцами асцидий они являлись излюбленным объектом для экспериментальной работы. Некоторые из этих яиц прозрачны, как стекло, другие же содержат пигментные зерна, движения которых служат прекрасными отличительными признаками, очень же ранняя диференцировка клеток дает возможность проследить их до конца. У этого объекта имеется только один недостаток, заключающийся в вязкости оболочки вокруг яйца, а также в наличии между поверхностью яйца и оболочкой внутреннего кольца особых клеток (рис. 18, а) и второго слоя {44} их над наружной поверхностью оболочки. Недавно был найден метод растворения оболочки без повреждения яйца.
 |
| Рис. 18. Движение веществ в яйце асцидий Styela во время оплодотворения (по Конклину). |
Зрелые яйца покидают яичник и собираются в яйцеводе. Полярное веретено в это время уже присутствует, но {45} полярные тельца не выделяются до тех пор, пока сперматозоид не проник в яйцо. Сперматозоид проникает в яйцо на вегетативном полюсе или очень близко от него. У Styela яйца содержат в поверхностном слое зерна желтого пигмента (рис. 18, а). Когда сперматозоид проникает в яйцо, внешний слой протоплазмы быстро скользит по поверхности, увлекая с собой окрашенные зерна (рис. 18, b). На одной стороне образуется желтый полумесяц, занимающий более полуокружности яйца (рис. 18, h—l). Этот полумесяц является первым указанием на билатеральный характер яйца.
Плоскость первого деления проходит через середину полумесяца (рис. 18 l), причем половина его попадает в одну клетку, а половина в другую. Попадает ли поверхностный протоплазматический слой и желтый пигмент полумесяца на предетерминированную сторону яйца или же их положение определяется точкой проникновения сперматозоида — до сих пор еще спорно. Однако одно, повидимому, достаточно хорошо установлено: головка сперматозоида двигается сначала в ту сторону, где находится середина полумесяца. Если сперматозоид проникает в яйцо не точно в вегетативном полюсе, а несколько в сторону от этой точки, а это, повидимому, является правилом, то эта эксцентрическая точка может служить детерминирующим фактором, приводящим к более экстенсивному накоплению поверхностной протоплазмы (и пигмента) на этой же самой или на противоположной стороне яйца. Если это рассуждение правильно, то билатеральное распределение содержимого яйца не предетерминировано, а является вторичным и, как и в некоторых других яйцах, обусловлено точкой проникновения сперматозоида.
Первое деление яйца проходит через анимальный полюс и через середину полумесяца (рис. 19, а—b), разделяя яйцо на две в точности равные части. Второе деление тоже проходит через полюс и под прямым углом к плоскости первого деления. Две из образовавшихся клеток несколько крупнее других двух и содержат большую часть полумесяца (рис. 19, с). Две меньших клетки содержат только рожки полумесяца и могут быть названы передними клетками. Протоплазма передних и задних клеток также несколько отличается по своему виду и, несомненно, их содержимое отличается и в других отношениях, а не только видимым количеством содержащегося в них пигмента или желтка. {46}
Борозда третьего деления проходит под прямым углом к первым двум и отрезает от каждой клетки (рис. 19, d) меньшую клетку на анимальном полюсе и более крупную на противоположном конце яйца. Две из последних содер-
 |
| Рис. 19. Стадии дробления яйца Styela (Cynthia) partita (по Конклину). |
жат большую часть желтой полосы полумесяца. Начиная с этого времени, деления продолжают происходить обычным образом, образуя клетки, которые могут быть идентифицированы по своему содержимому и местоположению. Судьба этих клеток будет описана в главе о гаструляции. {47}
Некоторые опыты сдавливания, проведенные на яйцах асцидий, показывают, что и эти яйца, подобно другим, делятся под прямым углом к сдавливающим пластинкам; но в отличие от сдавленных яиц морского ежа ни одно из яиц асцидий при освобождении от сдавливания не образует целиком нормального зародыша, хотя бы даже они и продолжали делиться. Некоторые из таких яиц дают начало зародышам с перемещенными органами. Это объясняется проще всего тем, что в этих очень рано диференцирующихся яйцах любое нарушение правильного распределения веществ приводит к катастрофическим последствиям.
Изучалось также влияние центрифугирования на эти яйца. Благодаря большей вязкости протоплазмы у таких яиц труднее вызвать слоистость, чем у многих других. Если яйца Styela центрифугируются перед оплодотворением, то желтый пигмент может быть сдвинут к одному полюсу; поверхностная протоплазма предположительно претерпевает свои нормальные движения и эти яйца могут образовать нормальных зародышей, причем пигмент может оказаться лежащим в любой части эмбриона. Это доказывает, что желтый пигмент как таковой не является органообразующим. Тем не менее его передвижение в нормальном яйце говорит об изменениях в перераспределении протоплазматического содержимого яйца, которые связаны с последующими влияниями на развитие. Имеются данные, показывающие, что у других асцидий, не имеющих пигмента, например, у Ciona, так же как и у Styela, происходят передвижения поверхностной протоплазмы, приводящие к образованию полумесяца.
Благодаря вязкой оболочке трудно получить фрагменты яйца асцидин, происходящие из определенных участков, и до последнего времени было невозможно отрезать фрагмент яйца из определенной области его. Тем не менее было показано, что фрагменты делятся и пытаются образовывать нормальные зародыши, но до тех пор, пока не будет точно определена связь фрагментов с определенной областью яйца, рискованно обобщать эти неадекватные результаты.
ДРОБЛЕНИЕ У ДРУГИХ ТИПОВ
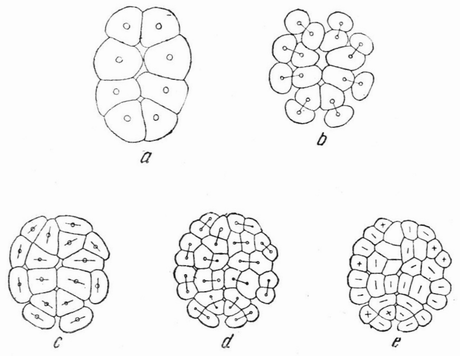
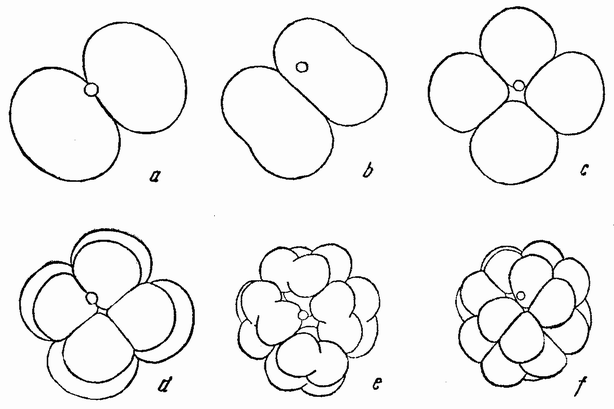
Хорошо известны и другие типы дробления, отличные от таковых у морского ежа и асцидий. Спиральный тип дробления аннелид и моллюсков отличается от предыдущего {48} типа главным образом тем, что при третьем дроблении вокруг полюса обособляются четыре клетки, микромеры, которые образуют вправо или влево закрученную спираль (рис. 20). При шестом дроблении на экваторе закладывается особая асимметричная клетка, называемая d4, дающая начало мезодерме или среднему слою зародыша
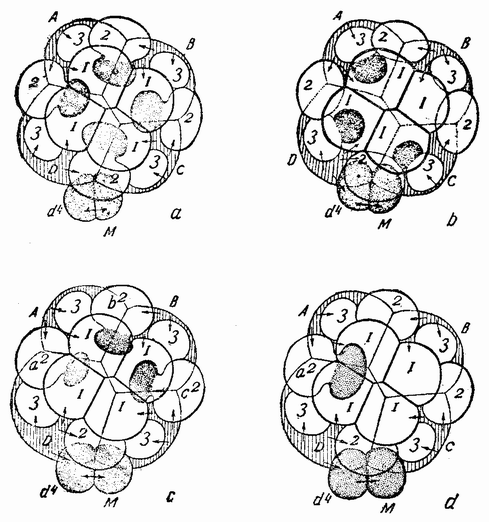 |
| Рис. 20. Поздние стадии дробления яйца планарии (a), аннелиды (b) и двух моллюсков (c и d). Показано положение клетки d 4 (no Вильсону). |
(рис. 20). Ее последующее деление на две маркирует первую резко различимую билатеральную плоскость зародыша.
Самки морской аннелиды Nereis всплывают иногда в летние ночи на поверхность моря и очень быстро плавают. Одновременно подымаются на поверхность и самцы. Если один из них приближается или сталкивается с самкой, то она немедленно выбрасывает все свои яйца, а самец одновременно изливает свою сперму. Яйца оплодотворяются {49} в морской воде. Каждое яйцо прозрачно и при выделении содержит крупное яйцевое ядро, которое быстро исчезает при оплодотворении, освобождая свои хромосомы. Яйцо окружено толстой оболочкой с радиальными порами. Под оболочкой имеется слой, содержащий жидкое вакуолизированное вещество, которое сразу проходит через оболочку, как только сперматозоид касается поверхности яйца.
 |
| Рис. 21. Дробление яйца Nereis. Образование первой (с) и второй (d) четверки микромеров. На рис. d видно деление первых микромеров (по Вильсону). |
Жидкость коагулируется в морской воде и становится желеобразной, покрывая собой яйцо. Она продолжает в течение нескольких часов набухать до тех пор, пока диаметр яйца вместе с окружающей его слизью не станет по крайней мере в три раза больше диаметра первоначального.
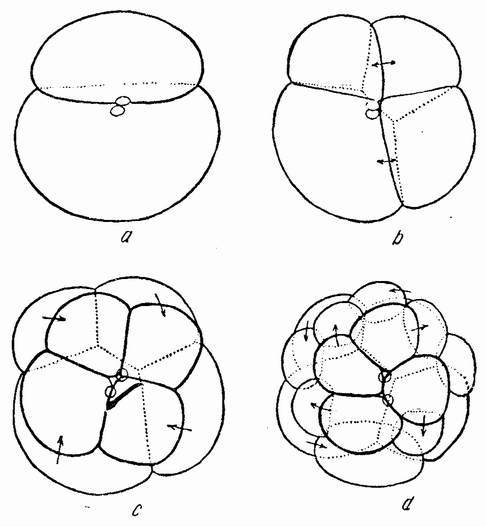
Проникновение сперматозоида через оболочку яйца продолжается более получаса. Его головка втягивается {50} выступающим бугорком оплодотворения; хвостик сперматозоида остается снаружи и служит маркером, с помощью которого может быть определено положение плоскости первого дробления по отношению к точке проникновения спермия. В большом проценте случаев они совпадают полностью и приблизительно совпадают в большинстве остальных случаев. Два полярных тельца выделяются внутри оболочки. Первое дробление (рис. 21, а) делит яйцо на несколько неравные части. Борозда второго дробления, проходящего под прямым уголом к первому, делит меньшую клетку на две равные, а большую на две неравные части (рис. 21, b). В результате вокруг оси яйца лежат три более мелких клетки приблизительно равной величины и одна более крупная клетка.
При третьем дроблении ясно обнаруживается спиральное деление яйца. Наискосок вокруг полюса обособляются четыре мелких клетки (рис. 21, с), каждая слева от своей сестринской клетки. Каждая клетка содержит очень прозрачную протоплазму, не имеющую желточных и жировых шаров, имеющихся в других клетках. Четыре микромера лежат в промежутках между четырьмя крупными клетками, причем из косого расположения веретен еще до того, как микромеры начинают отщепляться от крупных клеток, видно, что это расположение является не простым механическим заполнением полезного пространства после деления. При следующем, четвертом, дроблении (рис. 21, d) этот же самый процесс повторяется, но в это время веретена в крупных клетках поворачиваются в другом направлении, то есть вправо, давая начало второй четверке микромеров. Первые микромеры делятся, образуя спираль, закрученную в противоположную сторону. При пятом дроблении слева образуются еще четыре микромера — третья группа их. В это время справа добавляется четвертая группа микромеров из трех крупных клеток и четвертого микромера (d1), который является много более крупной клеткой (рис. 20, b), лежащей вдоль оси исходного второго деления. Эта клетка является предшественником большинства мезодермальных клеток зародыша. При ее делении на две равные клетки определяется средняя линия зародыша, проходящая между ними. Эта средняя линия не соответствует в точности какому-либо из первых двух делений, но очень близка к плоскости борозды второго деления. Микромеры продолжают делиться и, наконец, покрывают поверхность эмбриона. {51} Крупные клетки, содержащие желток, дают начало первичному кишечнику.
Деление яиц большинства моллюсков (рис. 20, c—d) за исключением каракатиц и осьминога, которые имеют крупные яйца, в принципе и часто даже в большинстве деталей, подобно делению яйца Nereis, несмотря на тот факт, что взрослые животные, принадлежащие к этим двум большим группам, совершенно различны. Все же сходство
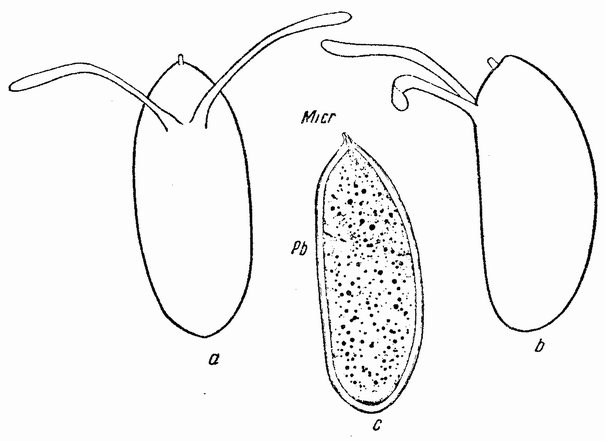 |
| Рис. 22. а — билатеральное яйцо дрозофилы с дорзальной стороны; b — с правой стороны; с — разрез яйца через среднюю плоскость и микропиле (Micr). Полярные тельца (Pb) выделились в верхней трети дорзальной стороны. |
самых первых стадий и тот факт, что происхождение их органов так сходно, вряд ли может оставлять сомнения в том, что эти две группы родственны и что в далеком прошлом их общие ископаемые предки, о которых мы не имеем никаких сведений, проходили через стадии развития, подобные обнаруживаемым у представителей обоих этих типов, живущих в наши дни.
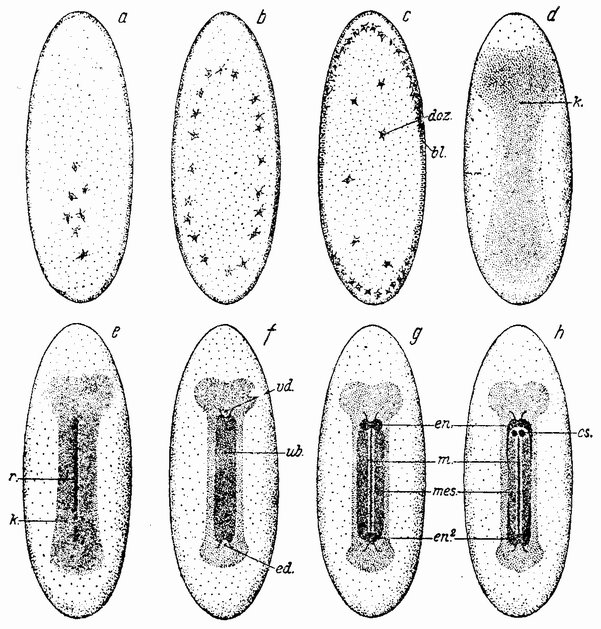
Яйца насекомых развиваются совершенно другим путем. Если предками насекомых были, как это принято думать, аннелиды, то они приобрели резко отличный тип {52} дробления. Яйца большинства насекомых имеют не сферическую форму, как это имеет место у большинства морских и пресноводных животных, а продолговатую, причем форма эта фиксируется на последних стадиях развития яиц и зависит от формы яйцевых трубок, в которых они образу-
 |
| Рис. 23. Дробление и образование зародыша у насекомого (по Мангольду). |
ются (рис. 22, а, b). Форма яиц обеспечивается очень твердой окружающей их оболочкой, которая пронизана одной или многими порами, находящимися не на конце яйца, а на одной из его сторон. Многие яйца билатеральны, причем одна сторона у них более плоская, чем другая (рис. 22, b—с). Стороны яйца соответствуют вентральной и дорзальной {53} сторонам эмбриона. Яйцо имеет билатеральную форму, когда оно готово к откладке. При продвижении яйца через заднюю часть яйцевода микропиле соприкасается с отверстием семяприемника, мимо которого оно проходит. В яйцо попадает несколько сперматозоидов, которые проникают в него вместе с хвостиками. Яйцо выделяет полярные тельца и яйцевое ядро глубже погружается в протоплазму. Ядра сперматозоидов движутся по направлению к ядру яйца и одно из них приходит в соприкосновение с последним. Тогда остальные ядра сперматозоидов пре-
 |
| Рис. 24. Первые четыре дробления яйца ланцетника (по Церфонтену). |
кращают двигаться к яйцевому ядру и впоследствии дегенерируют.
Внутренность яйца выполнена желточными гранулами, заключающимися в протоплазме; поверхность же яйца покрыта слоем протоплазмы, свободной от желтка (рис. 23, а). Веретено образуется поблизости от слившихся ядер и вокруг него собирается небольшое количество протоплазмы. Стенки ядра исчезают и хромосомы входят в веретено, где они делятся и расходятся к полюсам. Получается два ядра, однако протоплазма не делится. Эти ядра делятся опять, причем процесс этот продолжается до тех пор, пока число образовавшихся ядер не станет очень велико (рис. {54} 23, а—b). После каждого деления ядра расходятся в стороны и скоро образуют кольцо или даже сферу (рис. 23, b). По мере увеличения их числа, ядра приближаются к поверхности яйца и, наконец, оказываются лежащими в поверхностном слое протоплазмы (рис. 23, с). К этому времени имеется
 |
| Рис. 25. Каждая пара рисунков изображает противоположные стороны яйца лягушки во время первых дроблений. Более темная сторона на левых рисунках — так называемая вентральная сторона яйца, а более светлая сторона на правых рисунках — так называемая дорзальная сторона или сторона серого полумесяца. На рис. е и f изображены клетки дорзальной стороны, которые мельче таковых на противоположной стороне яйца на том же самом уровне (по Браше). |
уже несколько сот ядер. Далее, поверхностная протоплазма начинает дробиться, причем каждая борозда проходит между половинками делящегося ядра. Эти борозды располагаются радиально, но не доходят до внутренней стороны {55} наружного слоя протоплазмы, оставляя каждую клетку открытой на своем внутреннем конце; содержимое клетки и течение некоторого времени оказывается слитым с центральной желточной массой. На заднем конце яйца одно или несколько ядер переходят в особую протоплазму; эти клетки являются предшественниками половых клеток.
Яйца низших ракообразных заключены в твердую скорлупу. Они делятся на два, четыре, восемь и т. д. клеток, причем у речного рака и омара обнаруживают почти такой же тип дробления, как и у насекомых. У них имеется центральная желточная масса и поверхностный слой клеток, перегородки между которыми продолжаются до центра яйца.
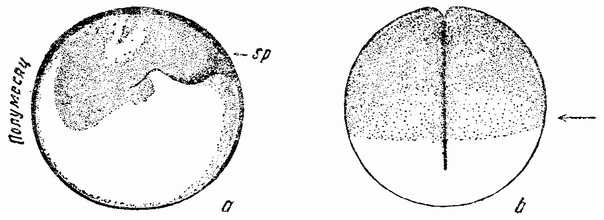 |
| Рис. 26. а — разрез через яйцо лягушки, через полумесяц в плоскости первого дробления. На стороне яйца, противоположной полумесяцу, виден след проникшего сперматозоида (sp); b — вид с поверхности на делящееся яйцо лягушки. Дробление разрезает полумесяц (по Шульце). |
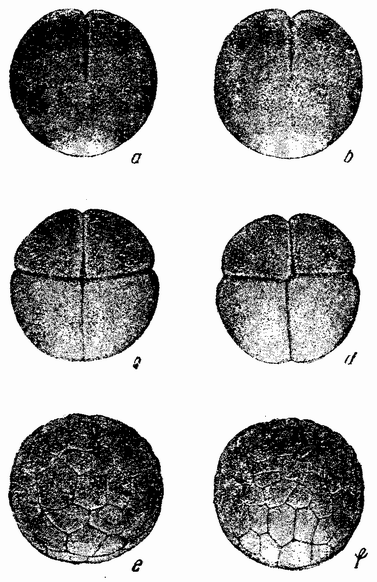
У позвоночных имеется два основных типа дробления. Ланцетник, являющийся низшим членом группы, имеет мелкие яйца, которые делятся сначала на две равные части, затем на четыре, восемь и т. д. (рис. 24, а—d), почти так же, как у асцидий. Яйцо лягушки, которое много крупнее, тоже имеет полное дробление (рис. 25, а—f). В первых стадиях развития этих яиц имеется одна особенность, которая напоминает развитие яйца асцидий. Как только в верхнее полушарие яйца проник сперматозоид, на той стороне яйца, которая лежит против точки проникновения сперматозоида, происходит движение поверхностного вещества. Серый полумесяц появляется вдоль края между черными и белым полушариями (рис. 26, a, b). Середина полумесяца, как и у асцидий, становится средней плоскостью зародыша. {56} Через его среднюю линию, как правило, проходит первое деление; но независимо от того, проходит ли это деление через середину полумесяца или нет, оно изредка может быть на 90° под углом к нему — средняя плоскость зародыша соответствует середине полумесяца.
Большинство яиц костистых рыб мельче, чем яйца лягушки, но тем не менее не имеют полного дробления. Их яйца состоят из центральной желточной массы, покрытой очень
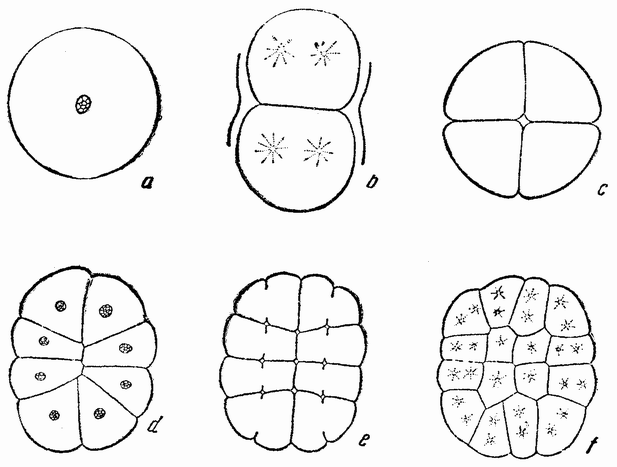 |
| Рис. 27. Первые четыре дробления яйца костистой рыбы (по Г. В. Вильсону). |
тонким слоем или шапкой протоплазмы, которая несколько толще над анимальным полюсом. У полюса через окружающую яйцо оболочку проходит небольшая трубка, по которой проникает сперматозоид. Первое дробление (рис. 27, b) состоит из прямой борозды в полярной шапке протоплазмы; борозда второго дробления проходит под прямым углом к первому; борозда третьего под прямым углом ко второму (рис. 27, с, d). Положение следующих делений несколько неправильно (рис. 27, е, f). Первая борозда сначала не сильно углубляется в яйцо; после {57} образования нескольких поверхностных бороздок самые центральные клетки делятся под прямым углом к последним, образуя внешнюю клетку и оставляя внутреннее ядро в поверхностном желтке.
Яйца рептилий и птиц имеют относительно огромную величину. Им свойственен желток, вокруг которого, по мере прохождения яйца по яйцеводу, откладывается слизь («белок»). Еще ниже в яйцеводе вокруг яйца образуется известковая скорлупа. Оплодотворение происходит, как только яйцо покидает яичник и входит в яйцевод. Дробле-
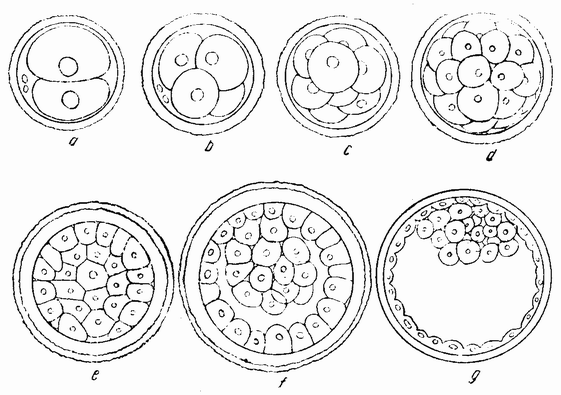 |
| Рис. 28. Сегментация яйца млекопитающего, f и g — начало образования бластоцисты и бластодермы (по Прентису). |
ние заключается сначала в образовании поверхностных бороздок, как и у яиц рыб. Образуется зародышевый диск, состоящий из многих клеток и лежащий на анимальном полюсе яйца.
Млекопитающие произошли от форм, имевших крупные яйца, подобные таковым рептилий. Низшее из живущих в настоящее время млекопитающих, Platypus, еще сохранило этот тип яйца, размеры которого три миллиметра в диаметре, но яйца всех других млекопитающих почти не имеют желтка, вследствие чего они очень мелки и едва видимы невооруженным глазом. Яйцо млекопитающего полностью делится на две, четыре, восемь клеток почти {58} равной величины (рис. 28, а—с). Внутри клеточной массы появляется жидкость (рис. 28, f), и клетки покрывают стенки образующегося пузырька (рис. 28, g), который постепенно увеличивается до значительной величины (рис. 29, a, b). В одной точке несколько раньше образовавшихся клеток соскальзывают под внешний слой, который затем покрывает
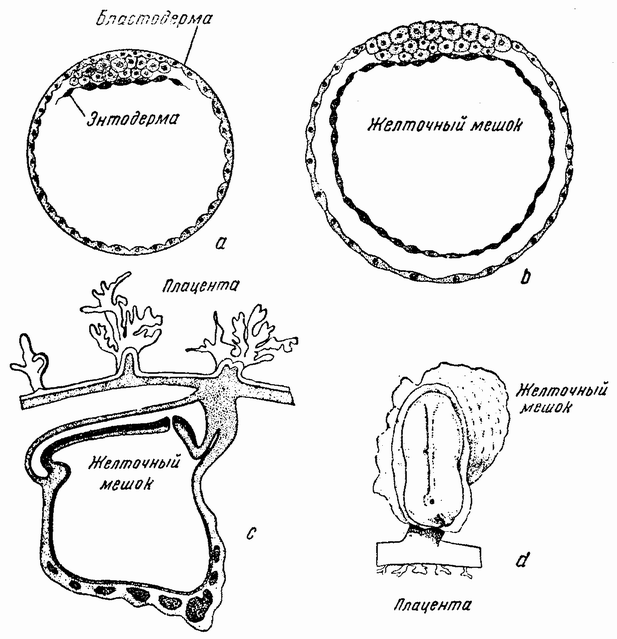 |
| Рис. 29. Развитие млекопитающего, а — бластоциста с бластодермой сверху; b — более поздняя стадия с эндодермой, образующей «желточный» мешок; с — зародыш более поздней стадии с «желточным» мешком и плацентой; d — вид молодого эмбриона с поверхности (из Кибель и Маль). |
их (рис. 29, а). Внутренняя группа клеток образует тогда небольшой диск, который затем дает начало эмбриону (рис. 29, d). Стенки пузырька соединяются со стенками {59} матки материнского организма (рис. 29, с), в которой зародыш претерпевает свое дальнейшее развитие.
В развитии млекопитающих имеются две стадии, которые казались бы непонятными, если не знать, что млекопитающие произошли из форм, имевших крупные яйца.
Молодые клетки, которые впоследствии станут яйцами, сначала лежат в поверхностном слое яичника (рис. 30, а).
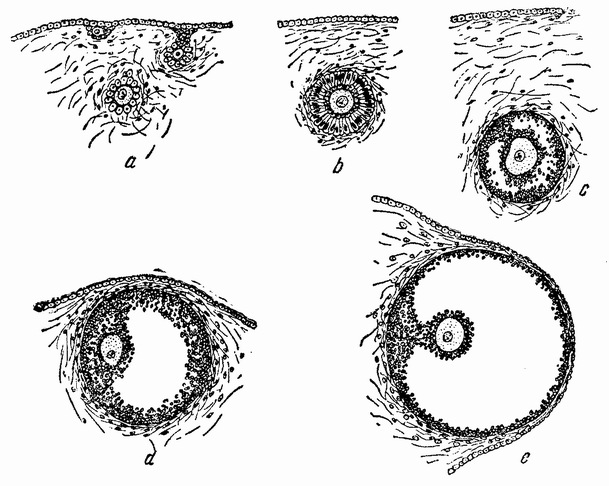 |
| Рис. 30. Четыре стадии яйца и граафова пузырька у млекопитающего (из Пэтена). |
Некоторые из них погружаются внутрь яичника, увлекая с собой группу других клеток, которые окружают каждое яйцо. По мере роста яйцевых клеток между клетками, непосредственно окружающими яйцо, и внешним слоем клеток появляется жидкое пространство (рис. 30 с, d, e); это пространство становится относительно огромным и на поверхности яичника выпячивается в виде шара, называемого граафовым фолликулом. Если бы не присутствие жидкости в фолликуле, то он напоминал бы крупный фолликул яичников птиц и ящериц, у которых яйцо, содержащее {60} желток, целиком заполняет внутренность фолликула. Если мы истолкуем величину фолликула как напоминание о состоянии предков, когда яйца млекопитающих содержали желток, то мы будем иметь своего рода объяснение современного состояния. Это объяснение постулирует, что яйцо млекопитающего сохранило в настоящее время некоторые из стадий развития своих предков.
Когда яйцо созревает, фолликул лопается. Воронкообразное отверстие яйцевода, открывающееся в полость тела, захватывает из яичника яйцо, которое проходит по переднему отделу трубы, и затем закрепляется в стенке
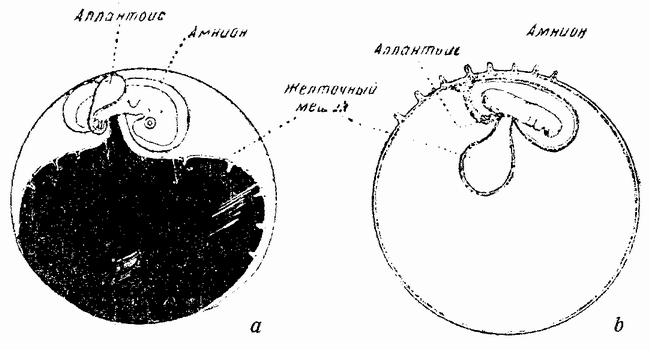 |
| Рис. 31. а — эмбрион и желточный мешок птицы; b — эмбрион и желточный мешок млекопитающего (а — по Лилли). |
матки, где и происходит его дальнейшее развитие.
Другая характерная стадия развития заключается в образовании желточного мешка, в котором отсутствует желток. Когда на бластодерме почти образовался зародыш (рис. 29, b), края энтодермы тянутся вокруг внутренней стенки эктодермального пузырька, образуя внутренний шар или мешок; этот мешок съеживается и отстает от наружной стенки и по мере вклинивания между ними мезодермы образует «желточный мешок» (рис. 31, b), основание которого связано со стенкой кишечника зародыша, и становится пупком. Впоследствии этот мешок втягивается в стенку пищеварительного тракта. Эмбрион, так сказать, «обманул» желточный мешок, хотя последний и не содержит в себе желтка. Другими словами, млекопитающее развивается так, как будто бы оно произошло из крупного, содержащего желток, яйца, подобного яйцу птицы (рис. 31, а) {61} или рептилии, которое следует еще способу развития своих предков. Желточный мешок является рудиментарным органом или эмбриональным атавизмом. Другими словами, генный комплекс, играющий роль в этой стадии развития, не был изменен. Конечно, проще предположить, что эти стадии зависят от особых генов, действие которых не затрагивалось изменениями в других генах, вызывавших дальнейшие эволюционные стадии.
| << | | {62} | | >> |