Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА V |
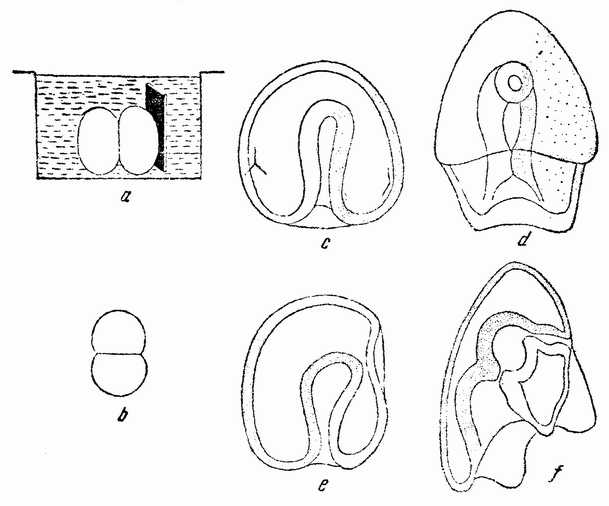 |
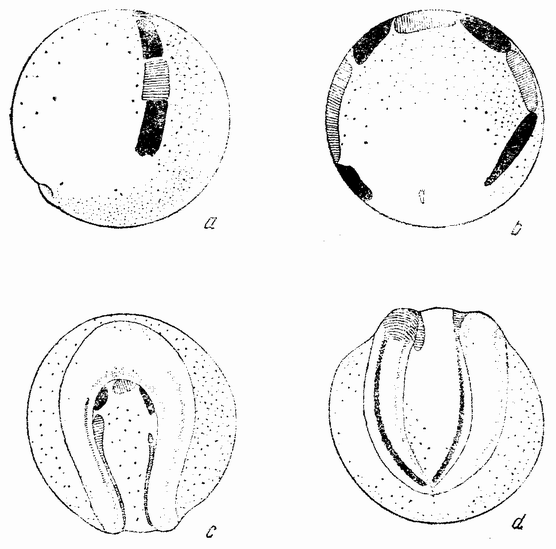
| Рис. 32. Способ окраски яйца морского ежа на двухклеточной стадии, а и b — с помощью кусочка агара, окрашенного «нильблау»; с—f — более поздние стадии гаструлы и молодой плутеус, развившиеся из таких яиц (по Убишу). |
агар-агара, которые были предварительно намочены в ниль-блау (рис. 32, а—b). Часть яйца, соприкасавшаяся с пластинкой, становилась синей. Когда один из бластомеров частично окрашивался, яйцо удалялось. Большинство развившихся эмбрионов было окрашено с одной стороны, показывая этим, что плоскость первого дробления совпадает с средней плоскостью зародыша. Однако в некоторых случаях окрашенной оказывается передняя или задняя {64} сторона эмбриона, что доказывает либо непостоянство этой связи, либо возможность изменения ее (рис. 32, с—f). У ланцетника процесс гаструляции (рис. 33, а—с) весьма сходен с гаструляцией морского ежа, за исключением лишь того, что инвагинированная часть (первичный кишечник) более широко открыта, образуя гаструлу, имею-
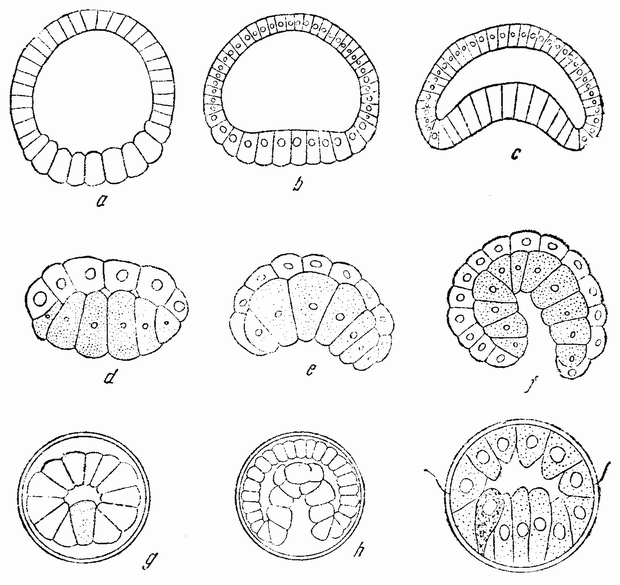 |
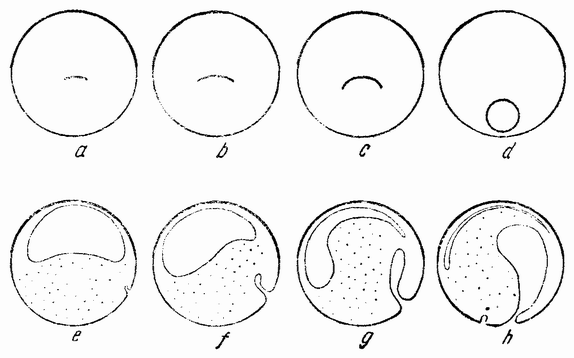
| Рис. 33. а, b, с — гаструляция у ланцетника; d, e, f — гаструляция у Clavelina; g, h — гаструляция у Lucifer: i — гаструляция у Eupomatus (по Румблеру). |
щую форму чаши. С окончанием инвагинации исчезает полость бластулы; отверстие гаструлы в дальнейшем редуцируется до небольшой поры на заднем конце, которая становится анальным отверстием. Дорзальная стенка первичного кишечника ущемляется, образуя спинную струну (рис. 34, А—В). С каждой стороны спинной струны {65} образуется ряд карманов. Эти выпячивания, отшнуровывающиеся из первичного кишечника, становятся полостью тела взрослого животного (рис. 34, С) и из их внутренних стенок образуются мускульные сомиты, составляющие основную массу взрослого животного.
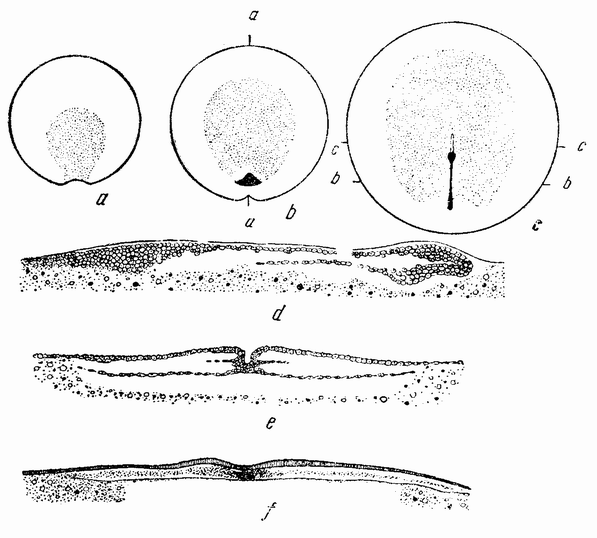
У лягушек и саламандр стадия бластулы (рис. 35, А) имеет над анимальным голушарием тонкую стенку, а внутри большую сегментационную полость, наполненную альбуминозной жидкостью. Нижнее полушарие состоит из крупных клеток, содержащих желток. Инвагинация начинается с одной стороны по близости от экватора, где
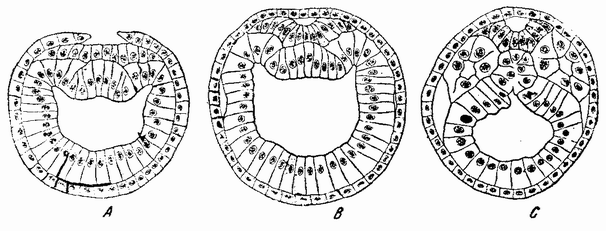 |
| Рис. 34. Поперечные разрезы через зародыш ланцетника, показывающие образование нейральной пластинки и трубки, спинной струны и кишечных мешков (по Церфонтену); |
располагается серый полумесяц (рис. 35, D). В этом случае инвагинация по своей форме скорее подобна щели, а не открытому рту, и происходит путем втягивания внутрь клеток наружной стенки в этой области. Поверхностные клетки спереди и с боков щели двигаются к ней и вворачиваются через ее край внутрь. Опыты маркировки внешних клеток краской (рис. 36) показывают, что обширный участок наружной поверхности перемещается внутрь, закручиваясь вокруг краев отверстия, и образует верхнюю и боковую стенки первичного кишечника. Край бластопора, имеющий форму полумесяца, движется по поверхности в нижнее полушарие (рис. 37) до вегетативного полюса, или еще дальше. Первоначальная внешняя стенка этого полушария становится дном первичного кишечника, а инвагинировавшиеся клетки становятся крышей. Во время этих перемещений происходят большие смещения клеток, особенно в верхнем полушарии. {66}
Из клеток, лежащих вдоль среднедорзальной стенки первичного кишечника, развивается спинная струна, а из клеток по обеим сторонам ее развивается мезодерма или средний слой, дающий начало мускульным сомитам, полости тела и т. п. И то, и другое образуется в основном тем же путем, что и у ланцетника (рис. 34), за исключением
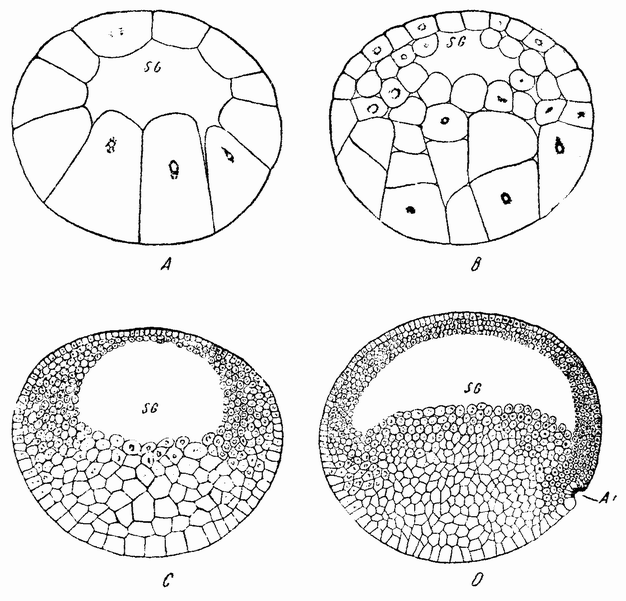 |
| Рис. 35. Разрезы через поздние стадии дробления и начало гаструляции яйца лягушки. |
лишь того, что в этом случае не происходит образования мешков. Рот образуется в той точке, где передний конец первичного кишечника приходит в соприкосновение с наружным слоем, который вворачивается внутрь и образует внутреннюю поверхность рта. Заднее отверстие первичного кишечника или отверстия гаструлы становится анусом. {67}
У позвоночных центральная нервная система ясно выражена с самого начала развития. Она появляется у эмбриона сейчас же после окончания процесса гаструляции. Слой поверхностных клеток, лежащих впереди бластопора, начинает сдвигаться вместе, образуя удлиненной формы нейральную пластинку. Края и передний конец пластинки
 |
| Рис. 36. а, b — стадии молодой гаструлы у тритона, на которую предварительно с помощью двух красок было нанесено полукольцо, охватывающее почти половину окружности яйца; с, d — последующее положение этих отмеченных участков (по Гертлеру). |
начинают приподыматься (рис. 36) и закручиваться, образуя полую трубку. Стенки этой трубки образуют мозг и нервный ствол, из которого вырастают нервные волокна ко всем частям тела. Нос и части органов зрения и слуха образуются инвагинацией поверхностного слоя клеток в головном участке. {68}
Зародыш птицы развивается из диска клеток на анимальном полюсе яйца, который со временем выталкивается над большой желточной массой. Первые следы гаструляции появляются на задней поверхности диска (рис. 38, а—b). Затем валик клеток, называемый первичной полоской, обозначает место бластопора (рис. 38, с), но прежде чем появляется первичная полоска, под диск углубляется пластинка клеток, дающая начало энтодерме (рис. 38, b, d).
У млекопитающих гаструляция происходит, повидимому, совсем таким же образом, как и у цыпленка, несмотря на
 |
| Рис. 37. Гаструляция у лягушки или тритона, а—d — вырост дорзальной губы бластопора; е—h медиальный разрез через те же стадии. |
то, что яйцо млекопитающего мельче и не содержит желтка. И все же оно развивается так, как будто бы в нем, как у ящериц и птиц, присутствует большое количество желтка. Диск клеток, из которого развивается эмбрион, лежит на поверхности большого пузырька (рис. 29, а). Периферические клетки диска простираются вокруг внутренней стенки пузырька (рис. 29, b), как будто бы для того, чтобы охватить, в действительности не существующую, большую массу желтка. Тем временем на бластодерме появляется первичная полоска (рис. 29, b) и из ее переднего конца происходит врастание клеток внутрь для образования спинной струны. Первичный кишечник и мезодерма образуются из нижнего слоя диска совсем таким же образом, как и у птиц. Процесс {69} гаструляции у млекопитающих дает много указаний на то, что он протекает по тому же пути, что у ящериц и птиц, но в то же время он выглядит как бы несколько сокращенным по сравнению с гаструляцией у последних. Не зная развития этих позвоночных, млекопитающие могут показаться одиноко стоящими в ряду остальных позвоночных,
 |
| Рис. 38. а, b, с — стадии развития бластодермы цыпленка; а — продольный разрез через b; поперечный разрез через первичную борозду с; f — поперечный разрез через спинную струну с (по Пэтену). |
зная же его, мы можем объяснить происходящие изменения предположением, что хотя зародыш млекопитающего и потерял свой желток, он все же сохранил тот способ гаструляции, которому следовали его предки. Обрастание несуществующей желточной массы для образования «желточного мешка» (рис. 31, b) происходит совсем так же, как у птиц (рис. 31, a). В конце концов этот мешок, как и у птиц, втягивается в первичный кишечник. Это является {70} характерным примером сохранения процедуры, свойственной предкам, но сейчас бесполезной — желточный мешок является рудиментарным органом.
Существовали многочисленные попытки найти механическое объяснение не только гаструляции, но и вообще явлению инвагинации, играющему важную роль в эмбриональном развитии. Эти объяснения основывались почти исключительно на аналогиях с механическими моделями и очень мало на результатах опытов с живыми яйцами. Однако основой для большинства гипотез послужил следующий подмеченный факт. Когда слой цилиндрических клеток инвагинируется для образования чаши, внутренние концы клеток становятся шире, а наружные уже (рис. 33, с). Другими словами, отдельные клетки принимают форму усеченного клина. Итак, если в одном участке эластичной наружной стенки бластулы клетки так изменят свою форму, что станут клинообразными, то произойдет автоматическое впячивание стенки внутрь. С другой стороны, если стенка была вдавлена внутрь снаружи или втянута изнутри, то клетки должны принять такую же самую форму, так как внутренняя поверхность становится больше наружной. Наблюдаемые изменения формы клеток будут в обоих случаях одинаковыми. Имеются, однако, некоторые соображения, которые, повидимому, исключают последнюю гипотезу. Так, например, не известно внешних сил, на которые можно было бы сослаться для объяснения вдавливания внутрь более толстой части стенки. Если бы такое воздействие имело место, то опустившимися клетками должны были бы быть более мелкие клетки крыши гаструлы. Если жидкость полости бластулы отсасывается, выдавливается наружу, или абсорбируется стенками, то опять-таки в результате этого опустится скорее крыша, а не более толстое основание бластулы.
Противоположная гипотеза предполагает, что более крупные клетки бластулы становятся шире либо потому, что они абсорбируют воду из полости бластулы, либо же потому, что на внутренней поверхности бластулы уменьшается поверхностное натяжение. С первого взгляда нельзя объяснить, почему набухание ограничено внутренней стороной клеток, т. е. почему не набухает вся клетка и не становится сферической. Конечно, внутренняя и наружные поверхности, вероятно, отличаются друг от друга. Казалось бы, что скорее внутренняя, а не наружная, часть должна {71} абсорбировать воду, но неясно, почему такое изменение должно иметь место не раньше, а именно в этот специфический период.
Конструировались различного рода модели, которые претерпевали инвагинацию, когда создавались такие условия, что в одной части стенки внутренняя поверхность ее абсорбирует жидкость изнутри скорее, чем наружная поверхность (рис. 39). В этих условиях происходит вворачи-
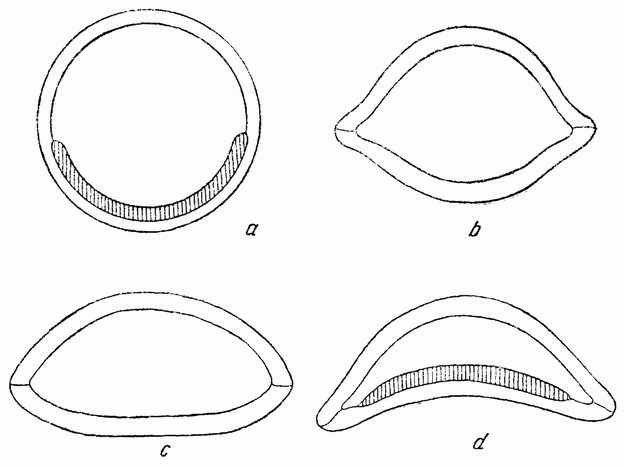 |
| Рис. 39. Модель из желатиновых полушарий для имитации гаструляции (по Шпеку). |
вание, но это еще не доказывает, что такого же рода различия существуют и между внутренней и наружной поверхностями стенки бластулы.
Пожалуй, наиболее правдоподобная гипотеза заключается в том, что инвагинация является результатом различий в поверхностном натяжении на внутренней поверхности стенки бластулы, обусловленных скоплением внутри углекислоты или каких-то других веществ. Как результат понижения поверхностного натяжения будет происходить перемещение клеточных веществ в ту сторону, где это {72} понижение происходит, причем внутренние концы клеток будут расширяться и инвагинация последует механически. Ни одна из этих гипотез в настоящее время не является удовлетворительной, потому что отстутствуют сведения относительно природы этих клеток; но они по крайней мере достаточны для того, чтобы считать, что Процесс гаструляции не исключает возможности физического объяснения. Как и в столь многих других случаях, имеющих место при развитии, анализ явления сводит его к поведению отдельных клеток; однако наблюдаемый факт, что активность всех клеток данного участка направлена к общей цели, побуждает нас искать такое объяснение этому явлению, которое трактует начало изменения как реакцию в ответ на внешние для клетки воздействия, а не как какие-то изменения, индивидуально происходящие в клетках. Но последняя альтернатива, однако, не была бы исключена, если бы удалось показать, что в этот определенный момент развития клетки данного участка индивидуально претерпевают внутреннее изменение, делающее их соответствующими физическим условиям системы в это время. Эти изменения, можно думать, являются кульминационной точкой тех изменений в протоплазме, которые начинаются в момент оплодотворения, или же они могут рассматриваться как новые изменения, которые вызываются в это время генами.
| << | | {73} | | >> |