Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА II |
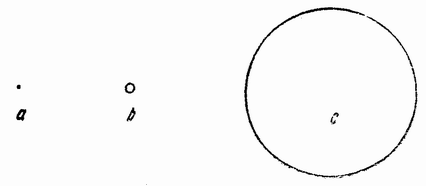 |
| Рис. 1. Диаграмма относительной величины яиц ланцетника (а), лягушки (b) и желтка куриного яйца (с). |
число и форма хромосом характерны для каждого вида. Способ, при помощи которого поддерживается это постоянство, стал понятен, когда Ван Бенеден (1883) сделал открытие, что ядро спермия привносит в яйцо половину типичного числа хромосом — отцовские хромосомы, и что ядро яйца перед оплодотворением тоже содержит в себе половинное число хромосом — материнские хромосомы. Другими словами, каждое зрелое яйцо и сперматозоид содержат ровно половину того числа хромосом, которое характерно для данного вида. Каким образом происходит редукция числа хромосом в зрелых половых клетках, было открыто позднее, в результате изучения изменений, имеющих место в яйцах и семенных клетках на последних стадиях их развития. Эти изменения будут описаны в следующей главе.
Анализ передачи по наследству родительских признаков показал, что хромосомы не являются конечными единицами наследственности, а содержат в себе более мелкие единицы, называемые генами. Последние расположены в {16} каждой хромосоме в линейном порядке, подобно бусам, нанизанным на нитку. Данные, что гены являются конечными наследственными единицами, покоятся не на прямом наблюдении, так как гены находятся за пределами видимости наших микроскопов, а на выводах из фактов наследственности.
К этому краткому обзору основных ступеней, приведших к современной концепции механизма наследственности, должно быть присовокуплено также и другое направление исследований, которое помогло выяснить роль хромосом в наследственности. Данные эти исходят из экспериментальной эмбриологии. Если яйцо разрезано перед оплодотворением на две части, то каждая из них может быть оплодотворена одним сперматозоидом и может развиться в зародыш. Одна часть яйца содержит ядро и ее развитие происходит так же, как и развитие целого яйца; она содержит после оплодотворения полное, диплоидное число хромосом. Другая же часть, вначале не имевшая ядра, после проникновения в нее спермия получает ядро сперматозоида с половинным числом хромосом, по одной каждого сорта. Это число хромосом является гаплоидным. И эта часть так же развивается и ее развитие устанавливает тот факт, что один набор хромосом достаточен для осуществления нормального развития. Правда, такие «гаплоидные» эмбрионы оказываются иногда слабыми. По крайней мерз в некоторых случаях было показано, что эмбрионы, начавшие свое развитие с половинным числом хромосом, достигнув взрослых стадий, удвоили число хромосом путем задержки одного из первых делений протоплазмы яйца. Тем не менее тот факт, что из гаплоидного куска может тоже развиться типичный эмбрион, показывает, что одного набора достаточно для образования характерных черт особи. В подтверждение этого вывода имеются также данные, полученные при изучении партеногенетического развития.
Другое доказательство значения хромосом было получено остроумным опытом Бовери. Было известно, что при проникновении в яйцо морского ежа двух сперматозоидов, яйцо при первом делении делится на три или четыре клетки вместо двух, как это происходит при нормальном развитии. Такие яйца не развиваются нормально. Как показал ранее Гертвиг, в таких диспермических яйцах первое деление хромосом происходит неправильно, и возникающие три или четыре клетки получают различные числа хромосом. {17} В то время было известно из результатов некоторых опытов Дриша, что если первые две клетки яйца морского ежа отделить друг от друга, то из каждой образуется нормальный эмбрион. То же самое имеет место при разделении четырех клеток. В этих случаях каждая клетка содержит, конечно, обычный двойной набор хромосом. Бовери же в противоположность этому нашел, что при разделении трех клеток диспермического яйца только одна из них, и то редко, развивается в нормальный зародыш (рис. 2). Он пришел к заключению, что неспособность большинства таких изолированных клеток нормально развиваться обусловлена неправильным распределением хромосом и что нормальный эмбрион развивается только из тех клеток, которые случайно получили по крайней мере один полный набор хромосом. Это заключение верно только отчасти. Одного набора хромосом действительно достаточно для развития, но если сверх него присутствуют также и другие хромосомы, то развитие оказывается нарушенным. В условиях опыта только редко может случиться, что в одну из трех клеток попадает в точности один набор хромосом и ничего больше. Так как превышение одного набора на любое число хромосом является нарушающим фактором, то для того, чтобы сделать достоверными цифровые отношения, на которых Бовери основывает свои выводы, необходим более подробный анализ способа распределения хромосом в таких яйцах. Позднее, генетическими опытами было показано, что основной набор может быть увеличен в три, четыре или большее число раз и тем не менее давать нормальное развитие, в то время как добавление к набору одной или двух хромосом может оказаться вредным.
Наиболее общее понятие о гене таково: это некая сущность с двумя основными особенностями. Во-первых, ген способен расти и делиться; во-вторых, он может вызывать изменения в протоплазме вне ядра — изменения, влияющие на химическую и физическую активность протоплазмы. Первое из этих свойств заключается в видимом делении хромосом, которые при каждом делении расщепляются продольно таким образом, что каждая половина содержит все свойства исходных хромосом. Другими словами, ген всегда делится, изменяя лишь свою массу, и затем растет, пока не достигает размеров исходного гена. Это же самое свойство должно быть у каждого саморазмножающегося нового гена. Способность деления является, следовательно, {18} свойством всей клетки, с тем лишь существенным различием, что при некоторых делениях яйца одна из дочерних клеток может содержать вещества, в некоторой степени отличные от таковых в ее сестринских клетках. Это неравенство первых делений яйцевой клетки может, как это устанавливается в последней главе, послужить нитью для пони-
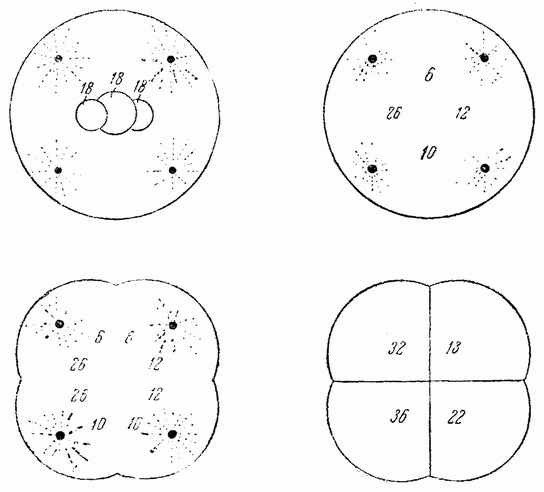 |
| Рис. 2. Схема диспермического яйца морского ежа с 3×18=54 хромосомами и четырьмя звездами (вверху слева). Остальные рисунки показывают один из многих возможных способов распределения хромосом по четырем образующимся клеткам (по Бовери). |
мания того, почему различные области сегментирующегося яйца развиваются различными путями.
Второе свойство гена не имеет прямого опытного обоснования, но покоится на предположительно логическом выводе из результатов генетического анализа. Эти генетические данные показывают, что когда ген мутирует, то он, не теряя своего свойства саморазмножения, вызывает изменения в признаке возникающей особи. Довод этот в действительности основан на обратной связи гена с {19} признаком, так, мы обнаруживаем сначала появление нового признака или группы признаков и затем уже при помощи анализа относим это изменение признака к изменению гена. Здесь важно то, что изменение может быть аналитически прослежено до отдельной точки или локуса в одной из хромосом, т. е. до единичного гена. Предполагая, следовательно, что мы правы в заключении, что изменение в признаке зависит от каких-то свойств нового гена, возникает вопрос о том, каким образом ген влияет на протоплазму клетки, ибо именно в протоплазме проявляется признак.
Можно предполагать, что эффект гена может обусловливаться каким-либо динамическим действием гена на окружающую протоплазму. Эта возможность не может быть в настоящее время доказана или опровергнута, но так как многие или даже большинство изменений в клетках по своей природе является химическими изменениями, то кажется более правдоподобным предположить, что ген выделяет какое-то химическое вещество, — может быть вроде катализатора,— которое вызывает некоторые химические изменения в протоплазме.
Прежде чем перейти к дальнейшему рассмотрению взаимоотношения генов и признаков, необходимо еще обсудить некоторые другие вопросы. Известно, что ген при мутации вызывает изменения во всем организме. Одни из этих изменений достаточно велики для того, чтобы быть вполне видимыми, другие мельче или даже настолько мелки, что ускользают от зрительного обнаружения, проявляясь только физиологически, вызывая смерть или изменение продолжительности жизни мутанта. На заре генетики, т. е. в начале этого столетия, предполагалось, что основой для генетической работы служат «самостоятельные признаки», откуда делался вывод, что каждый ген может вызывать специфический эффект одновременно только в одном признаке. Однако очень скоро было найдено, что этот преждевременный вывод является ошибочным, так как стал известным множественный эффект каждого генного изменения. Правда, в большинстве генетических работ выбирается отдельный признак, символизирующий ген, связанный с его проявлением, но этот выбор делается только потому, что либо этот признак один из наиболее легко отличимых, либо же потому, что он менее изменчив, т. е. менее подвержен влиянию внешней среды. {20}
Следующим пунктом, требующим обсуждения, является тот факт, что каждый признак взрослого организма есть продукт действия многих генов, или можно даже сказать всех генов, если проследить до яйца всю историю развития затронутого органа.
С этими вопросами тесно связано понятие генного равновесия. Выше уже отмечалось, что эмбриологические данные показывают, что для нормального развития необходим по крайней мере один набор генов; наиболее же обычным условием является наличие двух наборов. Но известно также много случаев, в которых четыре, шесть, восемь или больше наборов тоже давали нормальные результаты. Эти факты составляют часть данных, на которых основывается понятие генного равновесия,— понятие, которое первоначально, хотя и очень смутно, подразумевалось в приведенных эмбриологических данных. Однако оно приобрело значительно более определенный смысл, когда начали учитывать отдельные гены и в свете более новых данных экспериментальной генетики прежний умозрительный характер его уступил место более определенной формулировке.
Центральной мыслью теории генного баланса является положение, что на признаки действуют все гены и что то, что получается, является результатом их общего влияния. Если изменился, т. е. мутировал, только один ген, то продукт его в какой-то степени изменяется и некоторые органы оказываются затронутыми больше других; но тем не менее все гены играют при этом роль. Другими словами, новый ген действует только как дифференциал. Эта формулировка дает удовлетворительную картину конечных продуктов генов, но совершенно недостаточна для объяснения последовательности изменений, которые претерпевает эмбрион в течение своего развития.
| << | | {21} | | >> |