Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА VII |
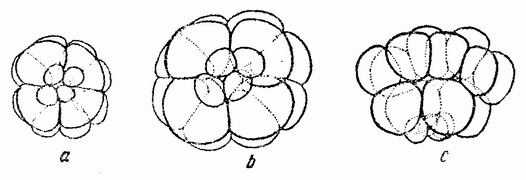 |
| Рис. 54. Шестнадцатиклеточная стадия вегетативной половины несегментированного яйца. Схема деления такая же, как и у целого яйца на соответствующей стадии (по Герсгадиусу). |
мера (на вегетативном полюсе), четыре макромера (в середине) и восемь мезомеров (на другом конце). Другими словами, она делится не как часть целого, а как целое яйцо. Пропорциональность между рядами клеток остается при этом такой же самой, как и в нормальном яйце (рис. 54, b).
Если же яйцо разрезано на две части по оси, т. е. под прямым углом к кольцу, то дробление протекает более изменчиво. Большинство половинок образует при этом нормальную шестнадцатиклеточную конфигурацию (рис. 54, с), особенно же если яйцо было разрезано перед оплодотворением, но некоторые половинки дают различного рода неравномерные типы дробления. Разница между различными типами дробления фрагментов может быть частично обусловлена трудностью разрезать яйцо точно через полюсы, различиями в закрывании поверхности разреза, наличием на одной стороне поверхности разреза {93} и т. д. Однако основные моменты совершенно ясны. Анимальное и вегетативное полушария яйца ведут себя не одинаково, причем это различие указывает на то, что они содержат различные вещества. Благодаря этому может быть в некоторой степени понятно пропорциональное развитие в вегетативных половинках и в меньшей степени в латеральных половинках, ибо митотические фигуры, определяющие плоскость дробления, регулируются общим количеством и распределением веществ яйца, из которых они развиваются. При половинном количестве вещества в половинном фрагменте величина митотической фигуры должна быть вдвое меньшей. Эта интерпретация сама по себе недостаточна для объяснения относительной величины бластомеров, ибо микромеры, например, в вегетативной половине, могут оказаться значительно крупнее, чем это ожидается. Должны быть, следовательно, допущены дополнительные факторы. И действительно, имеются данные, что вещество, попадающее в различные бластомеры, не обособлено. Например, не все вещество, попадающее в микромеры, содержится в нормальных микромерах, а часть его простирается дальше в анимальном направлении, так как для образования микромеров оно используется не полностью. Следовательно, для того чтобы дробление было пропорциональным, должно существовать некоторое восстановление или перераспределение близких слоев перед дроблением или во время него.
Было изучено поведение фрагментов неоплодотворенного яйца асцидии — Ascydiella. В настоящее время результаты этих опытов еще недостаточно ясны, для того чтобы послужить основанием для чего-то большего, чем только самых общих положений. Когда яйцо разрезано на две части по экватору или поблизости от него, то получаются различные типы ненормальных зародышей. В анимальной половине может нехватать энтодермы, а в вегетативной половине может быть нехватка эктодермальных частей. Могут быть получены парные зародыши, из которых один состоит из эктодермы, энтодермы, мезодермы и мезенхимы, а другой из эктодермы, энтодермы, нервной системы и спинной струны.
При разрезании яйца вдоль или почти вдоль его полярной оси может получиться пара половинных эмбрионов, два симметричных зародыша или же пара эмбрионов с дополняющими друг друга дефектами. Было предположено, {94} что половинные зародыши получаются в тех случаях, когда разрезы проходят через заранее детерминированную срединную плоскость, а симметричные эмбрионы, когда разрез перпендикулярен к этой плоскости. До тех пор, пока не будут описаны более тонкие детали стадий дробления фрагментов и раннее образование органов, такая интерпретация результата этих опытов останется очень неопределенной.
У немертины Cerebratulus развитие яйцевых фрагментов было изучено более подробно, чем у какого-либо другого
 |
| Рис. 55. Яйцо Cerebratulus во время стадий созревания, а — яйцевое ядро (зародышевый пузырек) еще присутствует, b — образовалось полярное веретено и с — выделилось первое полярное тельце (по Вильсону). |
животного. У этого объекта возможна точная локализация разреза и консистенция протоплазмы у него такова, что после операции фрагменты быстро округляются без видимых повреждений. Яйцо при откладке его самкой имеет в анимальном полушарии очень большой зародышевый пузырек (рис. 55, а), а на вегетативном полюсе удлинение студенистой оболочки и даже, в начале, короткий выступ вещества самого яйца.
Нормальное яйцо делится поровну на две, а затем на четыре клетки (рис. 56, а—b). Борозда третьего дробления образует почти равные клетки, но четыре анимальных клетки немного крупнее вегетативных (рис. 56, с). Это деление происходит спирально и против часовой стрелки. Последующие деления происходят попеременно по и против направления часовой стрелки (рис. 56, d). Позднее вегетативные клетки инвагинируют для образования гаструлы. Образуется типичная свободно плавающая личинка, называемая pilidium (рис. 57). {95}
После того как яйцевое ядро распалось для образования полярного веретена, яйцо может быть разрезано экваториально на две части и каждая часть оплодотворена. Анимальный фрагмент, содержащий яйцевой пронуклеус, выделяет два полярных тельца. Вегетативный фрагмент не выделяет ни одного полярного тельца, так как яйцевое ядро в нем отсутствует, но развивается только с пронуклеусом спермия, который служит ему «центром деления». Характер дробления обоих фрагментов во всех деталях
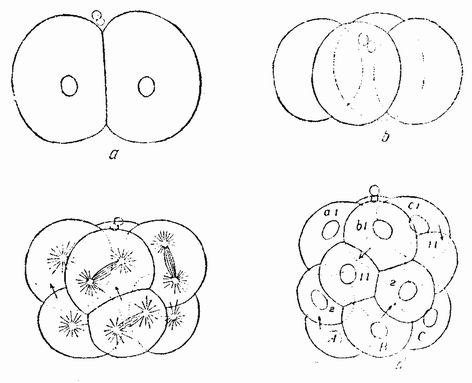 |
| Рис. 56. Нормальное дробление Cerebratulus (по Зеленому и Вильсону). |
подобен таковому целого яйца, включая чередование спиральных делений и относительные размеры бластомеров. Тот же самый результат получается независимо от того, проходит ли экваториальный разрез у экватора или выше или ниже него (рис. 58), за исключением лишь случаев, когда анимальный или вегетативный фрагменты исключительно малы.
Результаты практически одинаковы, когда яйцо разрезано на две части, меридионально, т. е. когда разрез проходит через ось, около нее, или ближе к одной стороне. Дробления части протекают при этом нормально, но могут быть и некоторые неправильности в делениях.
Были также изучены фрагменты, отрезанные во время образования полярных веретен, когда два пронуклеуса {96} конъюгируют и после того как начала появляться борозда первого дробления. Каждый из этих фрагментов дает начало небольшому нормальному зародышу. Тем не менее кажется, что по мере приближения момента первого дробления, удаление части яйца часто ведет к отклонению от типичного пути дробления и факты неравномерного дробления становятся более и более очевидными. Этого, конечно, и следует ожидать, так как изолированные бласто-
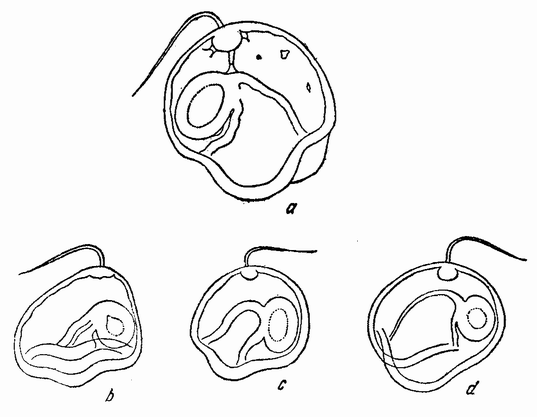 |
| Рис. 57. а — нормальная стадия пилидиума, у Cerebratulus; b, с, d — пилидии, образовавшиеся из фрагментов яйца (по Вильсону). |
меры этих, так же как и всех других, яиц дробятся неравномерно. Эти результаты подтверждают таким образом данные, полученные при изучении других яиц, а именно, что после оплодотворения в яйце происходит прогрессивный ряд изменений, предшествующих готовому начаться дроблению. В некоторых случаях известны изменения, вызывающие видимые сдвиги веществ яйца, которые связаны с последующими явлениями. Вряд ли можно сомневаться, что сами эти перемещения зависят от уже существующего распределения веществ яйца, но в какой степени они зависят от видимой слоистости яйцевых веществ или от невидимых физических различий в различных частях яйца — в настоящее время совершенно неизвестно. Однако {97} что развитие не зависит от строгой прелокализации органообразующих материалов в яйце как целом, ясно из того факта, что часть при определенных условиях проделывает то же, что и целое яйцо.
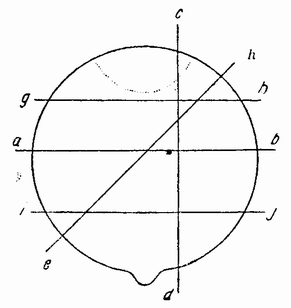 |
| Рис. 58. Схема плоскостей разрезов через яйцо Cerebratulus (по Вильсону). |
Изучались также фрагменты неоплодотворенных яиц некоторых других животных, как-то гребневика Beroё, аннелиды Chaetopterus, моллюска Dentalium, ланцетника и тритона, но так как они не дали существенно отличных результатов от только что приведенных примеров, то их можно не перечислять.
| << | | {98} | | >> |