Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА XV |
| | {187} | |
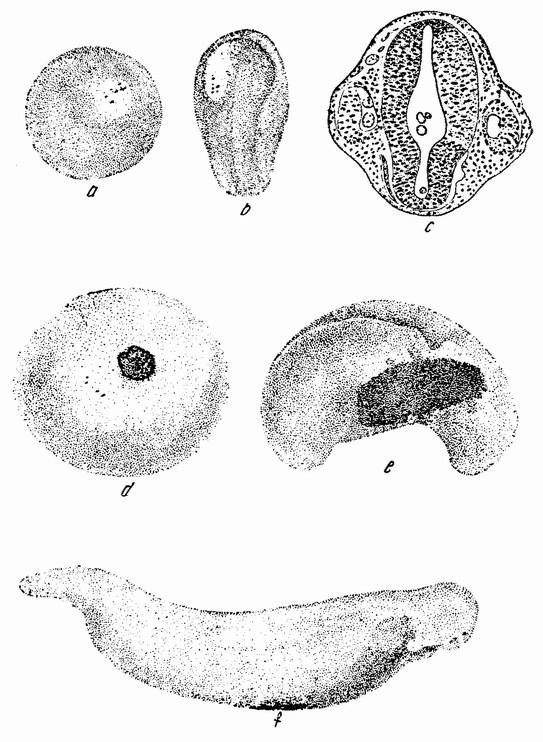 |
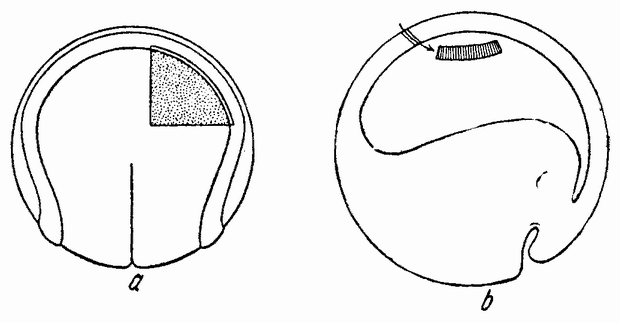
| Рис. 104. а — ранняя гаструла Triton taeniatus, в которую в ооласть проспективной нейральной пластинки пересажен маленький кусочек проспективной эктодермы Т. cristatus; b — более поздняя стадия а; с — поперечный разрез через зародыш, который развился из двух предыдущих стадий. Из имплантированного кусочка развилась правая сторона нейральной трубки и правый глазной пузырек; d — ранняя гаструла Т. cristatus, в которую в область проспективной эктодермы был пересажен кусочек вещества проспективной нейральной пластинки Т. taeniatus. На более поздних стадиях (е и f имплантант становится частью поверхностной эктодермы (по Шпеману). |
| | {188} | |
С другой стороны, если подобное изменение происходит на немного более поздней стадии, когда впереди бластопора обрисовывается нейральная пластинка, то кусочек нейральной пластинки, перенесенной на вентральную поверхность другого зародыша, погружается внутрь и образует пузырек, стенки которого диференцируются в нейральную трубку. Обратно, кусочек вентральной эктодермы, перенесенной на нейральную пластинку, не становится частью нейральной трубки. В последнем случае можно сказать, что диференциация зашла настолько далеко, что клетки не передиференцировываются и не принимают региональных черт своего нового местоположения. Эти опыты, хотя и не выясняют природу этих влияний, но они тем не менее показывают, что изменения прогрессивны и до некоторого момента могут быть обратимы. Даже если, как это постулирует теория мозаичности, в яйце и была предетерминация, то из опытов очевидно, что вначале более важен эффект положения.
Не менее важны и другие опыты, определяющие направление развития некоторых частей. Первый опыт этого рода показывает, что хрусталик глаза лягушки развивается в соответствии с наличием под эктодермой глазных пузырьков (выростов мозга). Это было показано двумя путями: во-первых, отрезанием конца глазного пузырька и имплантацией его под эктодерму соседнего участка на голове. Эктодерма вокруг имплантанта при этом утолщается и образует хрусталик; во-вторых, эктодерма участка, на котором должен был бы появиться хрусталик, удалялась и на обнаженный участок помещался новый кусочек эктодермы с другой части тела. Из этого кусочка тоже развивался хрусталик. Очевидно, хрусталик развивается в зависимости от наличия под ним глазного пузырька. Можно сказать поэтому, что хрусталик индуцируется в эктодерме присутствием оптической трубки: последняя является для хрусталика индуктором или, как выражаются в последнее время, организатором.
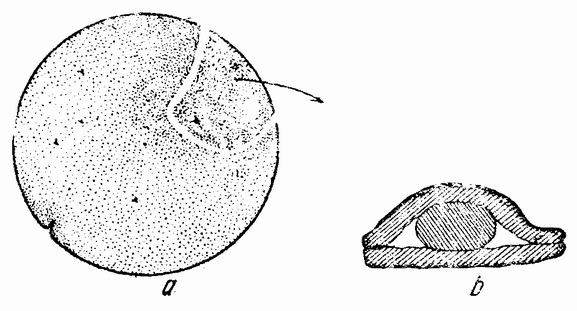
Аналогичные данные были получены при изучении развития центральной нервной системы тритона. Для того чтобы понять значение этих фактов, можно напомнить прелокализацию частей эмбриона у тритона, изображенную на рис. 105 А—Е. На рисунке А зародыш изображен сбоку, а на рис. В показан его вентральный вид сейчас же после образования дорзальной губы. Материал, {189} образующий нейральную пластинку, занимает верхний дорзальный квадрант, а будущая хордальная энтодерма и мезодерма лежат в квадранте под нейральной пластинкой. Последние участки ввернутся внутрь, обходя дорзальную губу бластопора. Судьба этих клеточных слоев была обнаружена путем маркирования краской различных мест на поверхности зародыша, благодаря чему име-
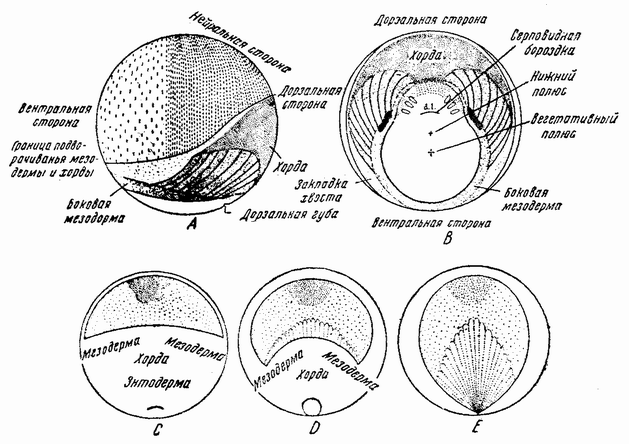 |
| Рис. 105. А и В — схемы участков поверхности зародыша тритона сейчас же после появления дорзальной губы гаструлы; А — вид сбоку; В — вид с желточного полюса (по Фогту). На трех нижних рисунках С, D и Е изображен участок проспективной нейральной пластинки (пунктир), хорды и мезодермы впереди дорзальной губы. На этих рисунках видно также удлинение материала нейральной пластинки во время закрывания бластопора, в то время когда хорда и мезодерма вворачиваются в губу бластопора (по Гертлеру). |
лась возможность проследить передвижения этих участков по мере последующего развития.
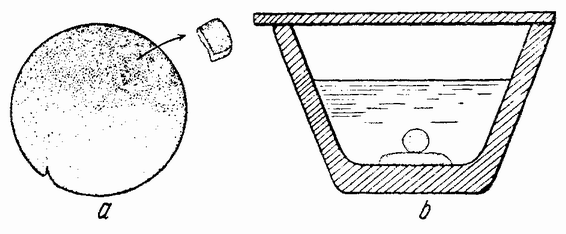
Самыми важными опытами были те, в которых вещество непосредственно спереди бластопора (рис. 106 а, проспективная хордальная энтодерма) и с боков его (проспективная мезодерма) было удалено и имплантировано на вентральную или латеральную поверхность другого, {190} более молодого зародыша, т. е. находящегося еще на стадии поздней бластулы или в начале гаструляции. Имплантант погружается при этом под поверхность, а затем вокруг и перед ним в эктодерме хозяина развивается неиральная складка (рис. 106 b). Могут даже появиться органы зрения, слуха и другие родственные структуры. Сам имплантант дает начало спинной струне и мезобластным сомитам. Про-
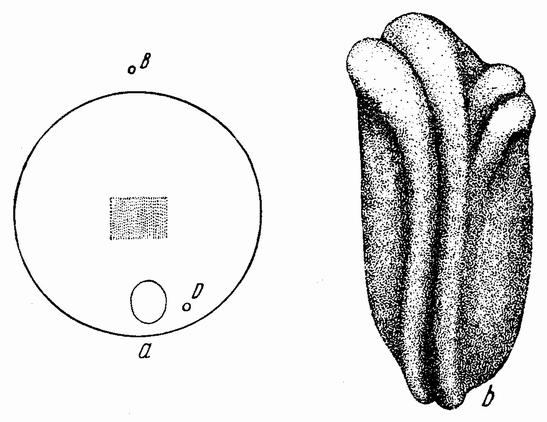 |
| Рис. 106. а — схема гаструлы тритона. Был удален кусочек поверхностной энтодермы нейральной пластинки этой нейрулы, а извлеченный кусочек дорзальной стенки первичного кишечника был введен в бластоцель молодой гаструлы. Эмбрион, развившийся из этой гаструлы, и его трансплантант изображены на рисунке b. Небольшая вторичная неиральная трубка находится сбоку переднего конца первичной нейральной трубки (по Гейнитцу). |
спективная хордальная мезодерма действует как индуктор, вызывая развитие в эктодерме нейральной трубки, которая в свою очередь может индуцировать в индиферентной эктодерме появление органов слуха, носовых ямок и других структур, т. е. органы, которые при нормальном развитии не появились бы в этих участках. Являются ли проспективная энтодерма, спинная струна или мезодермальные клетки стадии гаструлы равнопотенциальными, не вполне ясно из одного этого опыта, так как, как это изображено на рисунках, {191} между этими клетками на стадии гаструлы нет такой резкой разграничительной линии. Эти первые опыты дали основание предполагать, что индукция идет от клеток хордальной энтодермы, которая нормально вворачивается внутрь и становится крышей первичного кишечника, над которым у нормального эмбриона развивается нейральная пластинка. Наличие такого же рода реакции следовало бы предполагать и в опыте, в котором хордальная энтодерма оказывается лежащей где-либо под эктодермой. Однако этот простой и, казалось бы, логический вывод оказался слишком ограниченным для того, чтобы охватить все, что известно в этой области, ибо другие опыты показали, что индукция не является свойством одной хордальной энтодермы, а им обладают также другие клетки молодого зародыша.
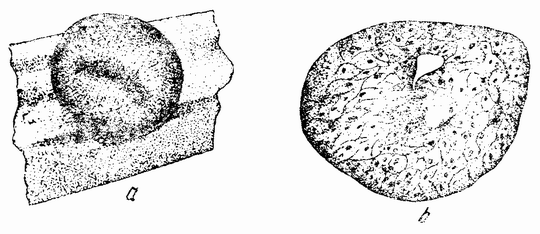
Следующие опыты были проделаны другим способом, а именно путем вставки кусочков из проспективной нейральной пластинки (взятых на различных стадиях) в бластоцель других эмбрионов, находящихся на стадии ранней гаструлы (рис. 107 и 108). Группа клеток в области проспективной нейральной пластинки была удалена с крыши ранней стадии гаструлы (в то время когда бластопор имеет U-образную форму) и помещена внутрь сегментационной полости молодой гаструлы (рис. 107 а, b). По мере протекания гаструляции введенный кусочек оказывается лежащим напротив крыши или краевой зоны реципиента, так как масса желточных клеток вдавливается внутрь и заполняет первичную сегментационную полость. Кусочек лежит, следовательно, между эктодермой снаружи и желтком или мезодермой изнутри. В общем можно сказать, что эктодермальный трансплантант сливается с поверхностной эктодермой хозяина и не действует как организатор, но в некоторых случаях индуцирует нейральную трубку. Из этого следует, что на этой стадии эти эктодермальные клетки, т. е. клетки проспективной нейральной пластинки, не всегда вызывают изменения в хозяине, как это делают подобные клетки, взятые из более поздней стадии.
В другой серии опытов из проспективного переднего конца поздней гаструлы был удален кусочек эктодермы с желточной пробкой (рис. 107 с) и перенесен в сегментационную полость более молодой гаструлы (рис. 107 d). Результаты были изменчивыми; иногда над прививкой появлялась только выпуклость, но в других случаях имелись {192} явные следы индукции, как это видно из развития структур нейральной пластинки (8 случаев из 29).
В третьей серии опытов был взят кусочек из переднего угла нейральной пластинки (рис. 108 a) и введен в сегментационную полость более молодой гаструлы (рис. 108 b); в этом случае над ним в «вентральной» эктодерме хозяина развивается нейральная пластинка. Кроме того, на перед-
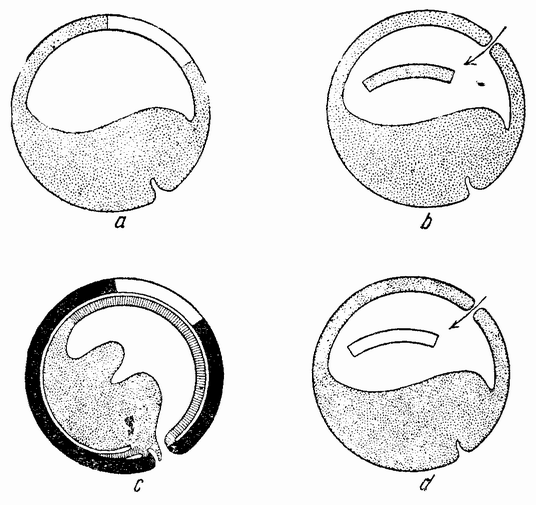 |
| Рис. 107. Схемы, иллюстрирующие трансплантацию частей гаструлы тритона, а — кусочек верхней стенки пересаженный в b; с — кусочек внешнего слоя верхней поверхности, трансплантированный в d (по Мангольду). |
нем конце нервной трубки могут развиться вторичные глазные бокалы, а из лежащей сверху эктодермы — хрусталик; могут также появиться слуховые пузыри и носовые ямки, которые иногда развиваются на переднем конце вторичного эмбриона.
В предыдущем опыте трансплантированный кусочек происходил из передней части нейральной пластинки. {193} Главными появлявшимися при этом органами были органы, характерные для переднего конца. Однако если частица взята из заднего конца нейральной пластинки более взрослого эмбриона, то эти передние органы отсутствуют и из тканей хозяина развивается хвостовая область. Имеются дальнейшие данные, что ответ хозяина в некоторой степени зависит от того, где расположен пересаженный кусочек, — ближе к передней области, дальше назад или в стороны. На результат влияет также стадия хозяина, на которой ему был привит кусочек. Другими словами, не только все части проспективной эктодермы хозяина
 |
| Рис. 108. Кусочек эктодермы из правого переднего конца нейралыгой пластинки зародыша тритона (а), трансплантированный в сегментационную полость молодой гаструлы (b) (по Мангольду). |
не одинаково отвечают на один и тот же индуктор, но и наоборот, влияние самих индукторов зависит от стадии зародыша или участка, из которого они происходят.
Новейшие опыты с организаторами у амфибий далеко отбросили виталистическое содержание термина организатор, которое могло бы вкладываться в него при выборе этого слова. Эти опыты показывают, что индукция может быть непосредственно обусловлена химической или физической реакцией между прививкой и хозяином, ибо эти эксперименты показали, что и мертвый трансплантант может вызвать индукцию, т. е. действовать как «организатор» или индуктор.
Если кусочки [проспективной эктодермы, нейральной пластинки или даже кусочки неоплодотворенного яйца убиваются нагреванием (до 60 °) и вводятся в бластоцель {194} молодой гаструлы, то они индуцируют развитие нейральной пластинки в проспективной эктодерме гаструлы. Путем вторичной индукции могут также развиться органы зрения, слуха и равновесия и т. д. Результаты индукции оказы-
 |
| Рис. 109. Кусочек верхней стенки молодой гаетрулы (а), помещенный между двумя лепестками живой эктодермы (b) (по Гольфтретеру). |
ваются совершенно такими же, как и при употреблении живых индукторов.
Такого же рода результат, хотя и не такой сильный, может быть получен другим путем. Если кусочек из молодой нейральной пластинки или дорзальной губы бластопора убивается нагреванием в солевом растворе и затем
 |
| Рис. 110. Кусочек поверхностного слоя молодой гаструлы из а, помещенный на мертвое вещество из другого яйца (b) по (Гольфтретеру). |
окружается двумя листками проспективной эктодермы, как это показано на рис. 109, то в последней развивается нейральная трубка, а позднее и характерная нейральная структура. Для протекания этого процесса существенно наличие индуктора, ибо если в солевой раствор помещаются {195} одни листки эктодермы, то они образуют только эктодерму. Индукция может быть вызвана также и другим образом. Частицы нейральной пластинки, предположительной эктодермы (рис. 110 а) или даже энтодермальные клетки убиваются нагреванием в солевом растворе. На них помещаются кусочки эктодермы (рис. 110 b), которые при соприкосновении с субстратом дольше чем один день развивают нейральные трубки (рис. 111 а—b). Эти резуль-
 |
| Рис. 111. а — образование ненральной трубки в кусочке, лежащем на убитом веществе, как на рис. 110; b — разрез через то же самое (по Гольфтретеру). |
таты показывают, что способность индуцировать нейральную трубку имеется и в мертвом веществе. Простейшее объяснение этого заключается в том, что индукция вызывается химической реакцией.
ВЛИЯНИЕ ХОЗЯИНА НА ПРИВИВКУ
Обратная реакция, а именно влияние хозяина на прививку, лучше обнаруживается в других опытах, особенно же при переносе молодых слуховых пузырьков и молодых зачатков конечностей в другие области зародыша.
Первые опыты касались слуховых пузырьков. Они появляются по одному с каждой стороны головы сначала в виде пластинки или чаши клеток из внутреннего слоя эктодермы, которая по мере ее погружения в лежащую под ней соединительную ткань закругляется в полый пузырек, из которого развиваются три полукружных канала, вестибулум, лабиринт и другие части внутреннего уха (рис. 112). На более поздних стадиях ухо является уже системой с тремя измерениями и имеет наружно-внутреннюю, дорзо-вентральную и передне-заднюю оси. {196}
Как только появился слуховой пузырек, покрывающая его эктодерма может быть приподнята и пузырек удален. Он может затем быть перемещен на то же место, но с повернутыми тем или иным образом осями, или транспланти-
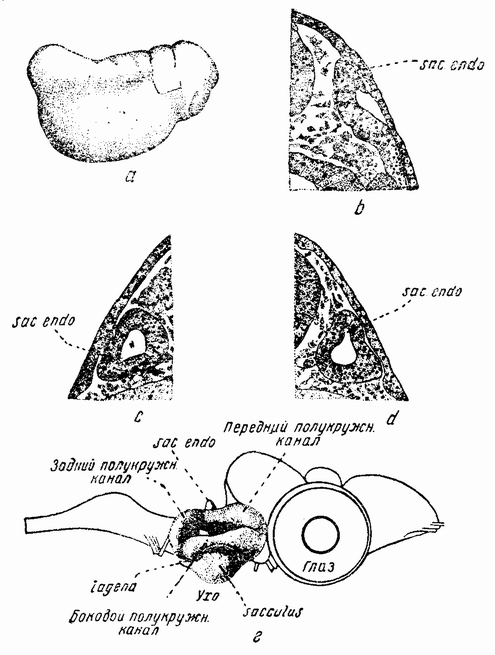 |
| Рис. 112. а — молодой зародыш лягушки; виден лоскуток эктодермы, приподнятый для того, чтобы обнажить ушной пузырек под ним; b — разрез через образующийся ушной пузырек; с — трансплантированный ушной пузырек, повернутый дорзо-вентрально; d — нормальный правый пузырек; е — полностью развитое нормальное внутреннее ухо (по Шпеману). |
рован на то же место, но с другой стороны эмбриона, повернутым или не повернутым, или же помещен где-либо в теле зародыша. Результат этих опытов несколько {197} затемнен, благодаря тому, что пузырек перед или во время его диференцировки поворачивается. Но в случаях, когда такого вращения не произошло, слуховой пузырек оказывается самодиференцирующимся, т. е. он развивается по своим собственным осям и не зависит от своего отношения к хозяину.
Сходные проблемы были изучены на зачатках конечностей амфибий, которые могут быть удалены и трансплан
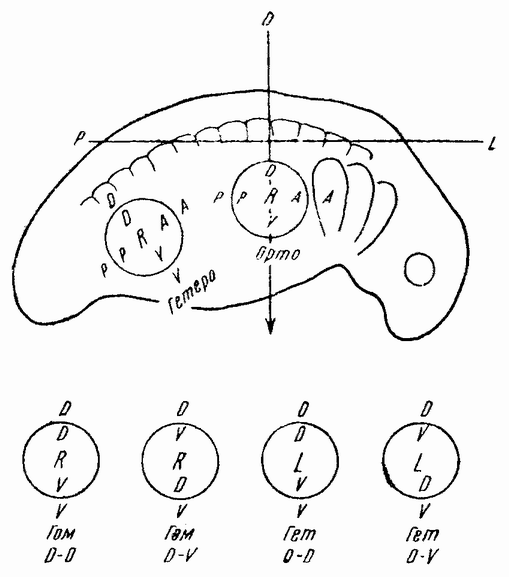 |
| Рис. 113. Схема, показывающая ориентировку привитых зачатков конечностей (по Гаррисону). |
тированы в том же положении с той же самой стороны зародыша или с противоположной с одной или двумя измененными осями, либо же перенесены на другие участки зародыша. Как показано на рис. 113, эти зачатки появляются в виде местных утолщений во внешнем слое мезодермы, которая образует сердцевину зачатка. Покрывающая эктодерма может быть приподнята и зачаток удален. Он может быть затем помещен под эктодерму в другом участке или с противоположной стороны другого эмбриона на место удаленного с этой стороны зачатка. Возможные {198} ориентировки для переднего и заднего зачатков показаны на рис. 113 (внизу). Кружком изображен на рисунке зачаток, а буквами внутри кружков его оси. Буквы вне кружков обозначают оси хозяина. Опуская детали, можно сказать, что в момент этих опытов зачаток конечности был «поляризован» только в одном передне-заднем направлении. Это значит, что в каком бы новом положении зачаток ни находился, его развитие находится в соответствии с этой господствующей осью, тогда как развитие в отношении двух других осей определяется его новым положением,
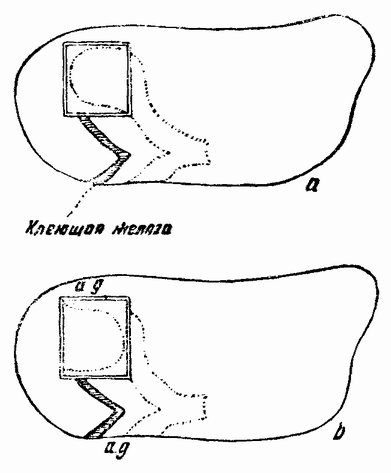 |
| Рис. 114. Схема, показывающая местоположение удаленного кусочка (а), повернутого на 180° и перемещенного, как показано на рис. 6. Клейкая железа служит для ориентировки. Пунктирная линия на обоих рисунках показывает очертания будущей жаберной крышки (по Экману). |
т. е. влиянием, исходящим от хозяина. Присутствуют ли во время пересадки и эти две последние поляризации и изменяются ли они последующими влияниями хозяина или же они еще не развились в зачатке — на этот вопрос этот опыт ответа не дает. Однако, повидимому, у некоторых видов осевые отношения могут быть фиксированными на более ранней стадии по сравнению с другими видами. Вероятно также, что у определенных видов дорзо-вентральная и латеральная оси тоже настолько детерминированы {199} на более поздних стадиях, что в новом положении их реверсия не будет иметь места.
Опыты с жаберными щелями лягушек показали, что впячивание эктодермы, которое встречается и сливается
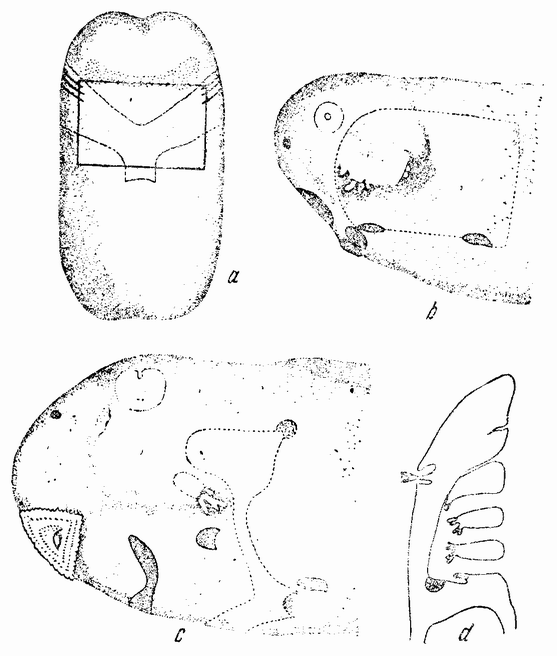 |
| Рис. 115. а — вентральный кусочек эктодермы, трансплантированный на обнаженный жаберный участок b. Как видно на рнс. d, в трансплантанте индуцированы жаберные щели; жаберный участок покрыт крышкой (с и d) (по Экману). |
с выпячиванием энтодермы глотки, индуцируется складкой энтодермы. Так, например, когда у молодого эмбриона удален квадратный кусочек эктодермы (рис. 114), который лежит в проспективной жаберной области, и пересажен, {200} будучи повернутым на 180°, на сходный участок другого зародыша, у которого предварительно был удален квадратный участок эктодермы такой же величины, то в прививке в эктодермальном выпячивании позднее появляются жаберные щели. Этот опыт не вполне убедителен, потому что кусочек повернут на 180°, что приводит вертикальное впячивание, хотя и в повернутом виде, но все же приблизительно, в то же самое положение, что и раньше. Кроме того, многое в опыте указывает на то что ко времени операции некоторая индукция уже имела место.
Более удовлетворительные результаты были получены при удалении кусочка эктодермы с вентральной поверхности над сердцем, как показано на рис. 115 а, и при пересадке его на обнаженную жаберную область другого эмбриона (рис. 115 b). Жаберные щели возникают в эктодерме прививки над лежащими под ними энтодермальными мешками. Позже на дугах появляются жабры (рис. 115 d). В придачу в трансплантированном кусочке появляется несколько мелких жабр на переднем краю прививки (рис. 115 с), которые могли быть индуцированными перед прививкой, так как боковые края вентрального кусочка эктодермы могут накрывать жаберную область. Но даже и в этом случае могла иметь место некоторая индукция,исходящая от хозяина, так как жабры не появлялись на заднем краю того же кусочка, лежащего позади жаберной области.
Еще более убедительны опыты удаления прямоугольного кусочка, простирающегося позади жаберной области в область головной почки, поворачивания его на 180° и имплантирования на тот же обнаженный участок зародыша, как показано на рис. 116. В этом случае область прививки, из которой образовались бы жаберные щели, лежит теперь позади жаберной области, как это показано вертикальными пунктирными линиями на рис. 116 a, тогда как эктодерма прививки, лежащая над будущими энтодермальными мешками, нормально лежит позади этого участка. Эктодермальные впячивания появляются позднее над энтодермальными мешками жаберной области и на них образуются жабры, как показано на горизонтальном разрезе на рис. 116 с. В этом случае имеется ясное доказательство индукции.
Будет ли эктодерма с любой части зародыша одинаково хорошо реагировать в таких опытах — сомнительно. В одном случае, в котором эктодерма из задней области была {201} помещена над жаберным участком, не развились ни щели, ни жабры. Здесь, как и в некоторых других опытах, имеются указания о наличии для каждого органа градиента в передне-заднем, дорзо-вентральном и, может быть, в радиальном направлении, который простирается кнаружи от проспективного центра, понижаясь с увеличением расстояния. Это, вероятно, может быть объяснено тем, что изменения в направлении будущего развития каждого участка про-
 |
| Рис. 116. а — кусочек поверхностной экзодермы над жаберным участком, достигающий уровня головной почки, удален и повернут в передне-заднем направлении; жаберные щели индуцируются в том конце прививки, который является теперь передним; прерывистые линии обозначают место, в котором должны были бы развиться жаберные щели; b — крышка, покрывающая жаберный район; с — на разрезе показаны индуцированные жаберные щели (по Экману). |
исходят, начиная с очень ранней стадии. Если эти изменения не зашли слишком далеко, то они могут исчезнуть или быть измененными в обратном направлении, если участок привит в новые условия окружающей среды. Сущность этих проспективных влияний в настоящее время совершенно неизвестна и в различных частях эмбриона она может быть весьма различной как по своему характеру, так и по количественному выражению. {202}
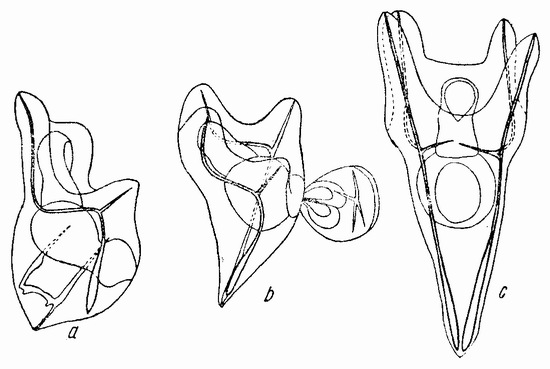
ОПЫТЫ ПО ПРИВИВКАМ С ЯЙЦАМИ МОРСКИХ ЕЖЕЙ
Различные комбинации половинок двух яиц морского ежа Paracentrotus могут быть получены следующим образом. После удаления яиц из их оболочки им дают развиваться до 16-клеточной стадии. Наличие микромеров на вегетативном полюсе дает затем возможность определить плоскость разделения групп бластомеров. Для того чтобы сделать это, яйца за десять минут до разделения, которое производится с помощью стеклянной иглы, помещаются в морскую воду, не содержащую кальция.
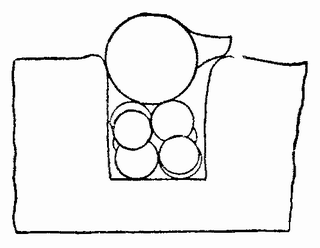 |
| Рис. 117. Схема, показывающая, как получается сращивание вместе бластомеров морского ежа (по Герстадиусу). |
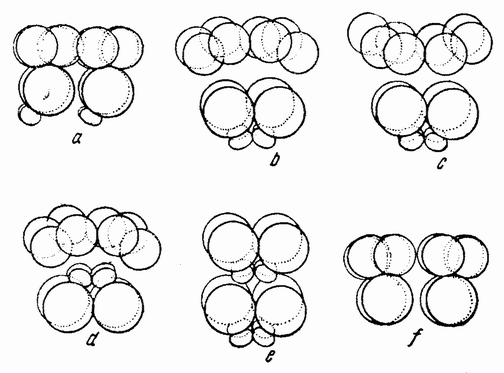
Когда полярность обеих соединяемых половинок одинакова, как показано на рис. 118 a, из них развивается нормальный плутеус. Так как разделение могло пройти либо через проспективную среднюю плоскость, если таковая в это время уже определена, либо под прямым углом к этой средней плоскости, то, повидимому, в это время такая плоскость отсутствует или, если присутствует, то может быть изменена и установлена заново. Когда половинки соединяются так, как показано на рис. 118 b, т. е. с обратной полярностью, то вегетативная область каждой половинки инвагинируется, как изображено на рис. 119 а и b. В этом случае образуется плутеус с двумя совершенно разделенными пищеварительными трактами, с двумя лежащими друг против друга анальными отверстиями, двумя средними и передними кишками и с одним общим ротовым отверстием (рис. 119 с). Скелет почти удвоен. {203}
Если половинки соединены, как изображено на рис. 118 с, т. е. ориентированы под прямым углом друг к другу, то гаструляция начинается в двух точках и два первичных кишечника раньше или позже соединяются, образуя раздвоенную или единую кишку. В этом случае развивается либо типичный, либо, более часто, ненормальный плутеус.
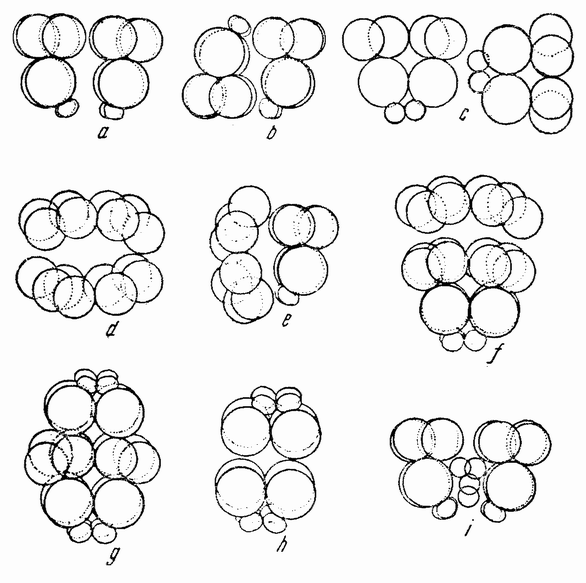 |
| Рис. 118. Схема различных способов соединения половинок 16-клеточных стадий морского ежа (по Герстадиусу). |
При соединении анимальных половинок, показанном на рис. 118 d, развивается бластула с двумя рядами ресничек, идущими в противоположных направлениях. Если соединены (рис. 118 е) меридиональная и анимальная половинки, то комбинация состоит из 12 мезомеров (эктодерма), 2 макромеров и 2 микромеров и развивается типичный плутеус (рис. 120 с). {204}
Если анимальная половинка присоединена к анимальному же концу целой 16-клеточной стадии (рис. 118 f), то такая комбинация содержит две трети эктодермы и одну
 |
| Рис. 119. а — гаструляция соединенных половинок (рис. 118); b — то же самое у другого подобного же соединения; с — развившийся двойной плутеус (по Герстадиусу). |
 |
| Рис. 120. а — атипичный плутеус, образовавшийся в результате соединения, изображенного на рис. 121 с; b — плутеус, развившийся из соединения изображенных на рис. 121 d; с — плутеус из комбинации е рис. 121 (по Герстадиусу). |
треть энтодермы, из нее может развиться плутеус, размеры которого будут крупнее нормального.
В случае соединения вегетативной половинки с анимальным концом целой 16-клеточной стадии (как показано на рис. 118 d) получается один плутеус с двумя кишечниками и одним ртом, сходный с изображенным на рис. 119 с. {205}
При соединении двух вегетативных половинок (рис. 118 h) происходит две инвагинации, причем личинки оказываются при этом совершенно ненормальными.
Четыре микромера имплантировались в другую 16-клеточную стадию (рис. 118 i), образованную из двух половинок, как изображено на рис. 118 а. Число мезенхимных клеток на стадии гасгрулы было удвоенным. Развивались типичные личинки без удвоения скелетных элементов.
В предыдущих случаях внутренние поверхности соединенных половинок были направлены друг к другу.
 |
| Рис. 121. Схема различных способов соединения половинок 16-клеточных стадий морских ежей (по Герстадиусу). |
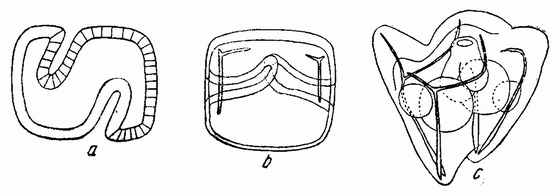
В другой серии опытов наружная поверхность одной половинки обращалась к внутренней поверхности другой половинки (рис. 121 а). Для того чтобы понять, как эта система выправляется, полезны будут следующие схемы. На рис. 122 а нарисована нормальная бластула, у которой вегетативный полюс заштрихован. Если эту половинку отрезать и повернуть так, чтобы ее поверхность расположилась напротив внутренней поверхности верхней половинки (рис. 122 b), то обе половинки изгибаются, как изображено на рис. 122 с. Затем две полости соединяются путем удаления перегородки (122 d) и получается единая полость (е—f). Внешние поверхности обеих {206} половинок находятся теперь на внешней поверхности одной бластулы, но здесь имеется два центра вегетативных клеток, лежащих на экваторе друг против друга.
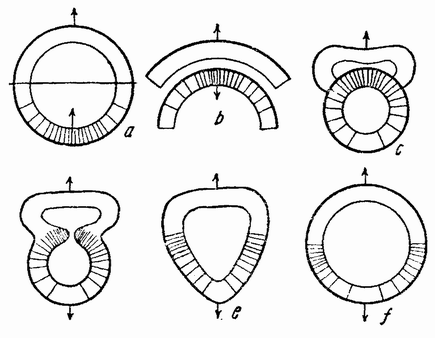 |
| Рис. 122. Схемы, показывающие результат соединения вегетативных половинок, повернутых и соединенных с анимальной половинкой так, как показано на рис. b (по Герстадиусу). |
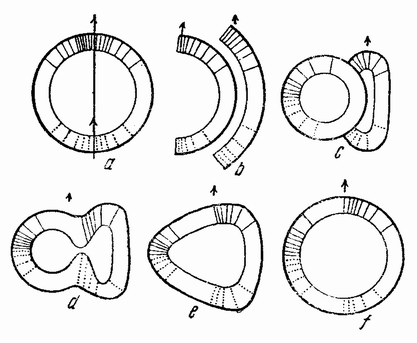 |
| Рис. 123. Схема, показывающая результат соединения меридиональных половинок 16-клеточных стадий, как показано на рис. b (по Герстадиусу). |
На рис. 123 а представлены две разделенные по анимальной оси половинки, одна из которых перевернута и {207} ее внешняя поверхность помещена внутрь другой половинки (123 b). Обе половинки закругляются, как показано на рис. 123 с. Перегородка удаляется (d) и получается единая полая бластула. Вегетативная область находится на той же стороне бластулы, но несколько в сторону от нее (d—f).
С помощью этих схем легко понять поведение соединенных половинок 16-клеточной стадии. Если две половинки соединены, как на рис. 121 а, получаются как типичные, так и атипичные плутеусы. Если две половинки соединены, как на рис. 121 с, то развиваются тоже и типичные и атипичные плутеусы. При обратной комбинации (рис. 121 d) развитие идет совершенно неправильно (рис. 120 b). Некоторая регуляция в этих случаях происходит, но гаструляция нарушается и плутеусы оказываются ненормальными в отношении энтодермы (рис. 120 b).
Если восемь клеток двух вегетативных полушарий соединяются с той же ориентировкой (рис. 121 b), то происходит две инвагинации, а получающиеся плутеусы оказываются очень ненормальными.
Если с каждой из двух половинок удаляются микромеры и половинки соединяются затем так, как изображено на рис. 121 f, то не образуется первичной мезенхимы, а с опозданием появляется вторичная мезенхима. В этом случае может развиться нормальный плутеус.
Эти опыты прививок показывают, что верхняя и нижняя половинки уже специфичны в отношении их будущего развития. Этого можно было ожидать из поведения анимальной и вегетативной половинок при изолировании их, а также и из поведения анимальных и вегетативных половинок несегментированных яиц. Существует некоторое взаимное восстановление клеток, но в целом при наличии двух групп макромеров очевидна тенденция удвоения (что дает начало первичному кишечнику). В какой степени макромеры действуют в качестве индуктора для остального эмбриона, не вполне ясно. Факты говорят скорее противоположное.
| << | | {208} | | >> |