Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА XIII |
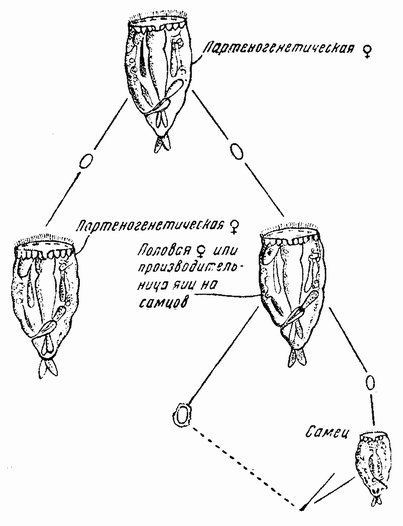 |
| Рис. 89. Жизненный цикл Hydatina senta. |
которого заполнена главным образом семенными клетками. Во-вторых, если эта же самка была оплодотворена спермой самца вскоре после того как она вылупилась из яйца, то сперматозоиды находят себе путь к молодым яйцевым клеткам и проникают по одному в каждую из них. Эти яйца вырастают тогда до более крупных размеров, чем не оплодотворенные яйца на самцов и покрываются скорлупой. Каждое яйцо выделяет два полярных тельца и во время одного из двух полярных делений число хромосом уменьшается {158} вдвое. Затем ядра яйца и спермия соединяются вместе для образования сегментационного ядра, которое, следовательно, имеет диплоидное число хромосом. Этот эмбрион всегда развивается в самку, которая называется самкой-основательницей и от нее начинается новый партеногенетический цикл.
Существует другой важный факт, касающийся стадий созревания семенных клеток гаплоидного самца. Повидимому, образуются два сорта сперматозоидов, из которых функционирующим является только один из них. Каждая зрелая семенная материнская клетка, являющаяся гаплоидной, делится один раз, причем все хромосомы расщепляются на две половинки, расходящиеся к противоположным полюсам; каждая клетка до сих пор содержит гаплоидное число хромосом. Одни из этих клеток, не претерпевая второго деления, становятся функционирующими сперматозоидами, другие же или некоторые из последних делятся еще раз и дают начало нефункционирующим сперматозоидам. Можно, следовательно, сказать, что единственными функционирующими сперматозоидами являются спермин дающие самок, но имеется ли здесь механизм половых хромосом и если имеется, то как эти хромосомы разделяются, неизвестно.
Одной из наиболее интересных особенностей в жизненном цикле коловраток является то, что внешние условия могут так влиять на партеногенетическую линию, что в ней начинают появляться половые формы. Влияние внешних условий изучалось с разных сторон и, как это было описано выше, было исследовано несколько агентов так называемых производителей-самцов для повышения числа особей, дающих начало самцам. В число этих агентов входят температура, кислород, пища и аккумуляция побочных продуктов обмена по мере старения или перенаселения культуры. Было с убедительностью показано, что из всех этих агентов эффективен только один, а именно пища. Так, например, если Hydatina питается бесцветными жгутиковыми инфузориями (Polytoma), то она существует неопределенно долгое время как партеногенетическая линия; но если в какой-либо момент в культуру будут в изобилии введены зеленые жгутиковые, Chlamydomonas, то почти все следующее поколение самок образует яйца на самцов, а если самки были оплодотворены, то половые яйца. Окончание партеногенетической линии и начало {159} половой фазы обусловлены в этом случае изменением пищи. Как установлено выше, та же самая особь, которая образует яйца на самцов, при раннем оплодотворении, образует также и половые яйца. Можно сказать, что внешняя среда создает пол, но не определяет его. Определяется же пол присутствием или отсутствием сперматозоида.
У одной группы ракообразных, у Cladocera, обнаруживается некоторое сходство с коловратками в том отношении, что у них может быть длинный ряд диплоидных пар-
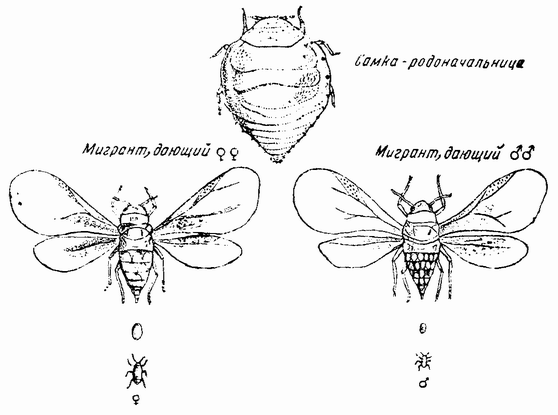 |
| Рис. 90. Жизненный цикл Phylloxera caryecaulis. |
теногенетических самок, яйца которых выделяют только одно полярное тельце. Как результат изменения во внешних условиях могут появиться самки, дающие начало самцам или половым яйцам, но условия, определяющие тип яйца, в этом случае являются, повидимому, не теми что у коловраток. Например, самки Cladocera, дающие самцов, появляются при перенаселении культуры, а у коловраток, наоборот, перенаселение препятствует появлению таких самок. Предполагалось, что у Cladocera аккумуляция продуктов выделения самих животных (или присутствующих бактерий) вызывает повышенное образование самцов. Прибавление мочевой кислоты, хлористого этана или углекислоты увеличивает процент самцов. Данных о {160} том, что эти агенты вызывают изменения и у коловраток не имеется, наоборот, в застаревших культурах на конском навозе процент самок производящих самцов уменьшается.
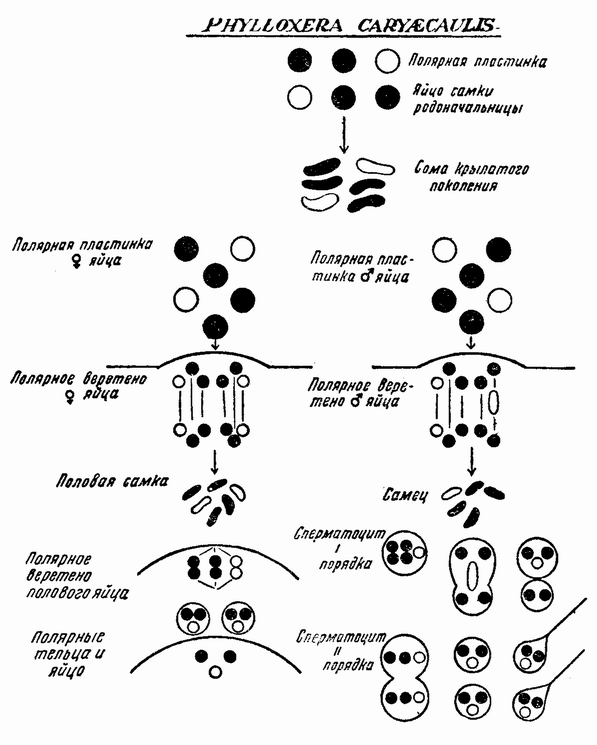 |
| Рис. 91. Хромосомный цикл P. caryecaulis. |
У насекомых чередование партеногенетического и полового поколений имеют также тли и филоксеры (рис. 90). У первых было показано, что длина дня, т. е. светлых часов, является одним из агентов, вызывающих изменение, но {161} неясно, действует ли этот фактор непосредственно или каким-то косвенным путем. В этих двух группах известны два других факта, объясняющих некоторые отношения, не обнаруженные у коловраток (l). Партеногенетические самки имеют диплоидное число хромосом и выделяют одно полярное тельце (рис. 91). В конце партеногенетической линии появляется самка, яйца которой тоже выделяют только одно полярное тельце, но, когда все другие хромосомы делятся, одна из Х-хромосом целиком отходит в полярное тельце, оставляя своего партнера — другую половую хромосому — позади (рис. 91, справа). Это яйцо, следовательно, дает начало самцу, который имеет диплоидное число хромосом минус один Х(2). Когда семенные клетки у этого самца претерпевают первое деление созревания, то каждая из хромосом коньюгирует со своим партнером за исключением лишь Х-хромосомы, которая не имеет партнера. Члены каждой пары движутся затем к противоположным полюсам, а единственная Х-хромосома к одному из полюсов (рис. 91). Клетка, получающая Х-хромосому, становится крупнее другой и две дочерних клетки разделяются. Одна из них имеет гаплоидный набор (включая X), другая же содержит гаплоидный набор минус Х-хромосому. Крупная клетка претерпевает затем второе деление созревания, при котором каждая хромосома, также как и Х-хромосома, расщепляется. Каждая клетка становится функционирующим спермием на самку, так как она содержит Х-хромосому. Меньшая же клетка, являющаяся продуктом первого деления, дегенерирует.
У других самок, появляющихся одновременно или на поколение позже, чем самки, дающие самцов, развиваются половые яйца. Они выделяют два полярных тельца и сохраняют гаплоидное число хромосом, в числе которых имеется одна Х-хромосома (рис. 91, слева). При оплодотворении спермием на самку, восстанавливается полное число хромосом (2А+2Х). Из этого яйца, называемого покоящимся яйцом, развивается самка-основательница, являющаяся родоначальницей новой линии партеногенетических самок.
Здесь половые хромосомы опять играют свою обычную роль в определении пола, причем фактором, изменяющим партеногенетическую фазу в половую, является внешняя среда.
Существуют различные виды животных, представленные в некоторых географических областях только самками, {162} а в других и самками и самцами. Первые размножаются путем партеногенеза; вторые — из оплодотворенных яиц. Например: один из равноногих раков, Trichoniscus, представлен на севере Европы только самками, которые являются триплоидными и содержат 24 хромосомы; на юге же найдены диплоидные самцы и самки, имеющие 16 хромосом. Если яйца этих половых самок не будут оплодотворены, то они не развиваются. Здесь, следовательно, имеется две определенных расы одного и того же вида, причем, так как они не могут скрещиваться между собой, можно сказать, что это два вида, которые идентичны во всем, кроме способа их размножения.
У другого ракообразного, Arthemia salina, имеются тетраплоидные партеногенетические расы с 84 хромосомами и диплоидные половые расы с 42 хромосомами. Среди кузнечиков известны также случаи, когда яйца при оплодотворении дают начало и самцам и самкам, причем пол особей регулируется двумя сортами сперматозоидов, а без оплодотворения образуют только самок. Некоторые виды Phasma размножаются только партеногенетически; другие же виды имеют половое размножение. Можно было бы привести много других случаев, в которых партеногенез занимает определенное место в жизненном цикле организма.
Эти примеры, число которых может быть увеличено, служат для демонстрации распространенности партеногенеза в животном царстве. Однако это явление известно также и у многих растений. Эти случаи подчеркивают тот факт, что способность к развитию заключается в самих яйцах. Получается, следовательно, картина, отличающаяся от обычного представления, подчеркивающего, что начало развития является результатом оплодотворения. Я думаю, что правильнее будет сказать, что главная функция проникновения спермия в яйцо заключается в удалении препятствия, удерживающего яйцо от развития. С этой точки зрения партеногенетическое яйцо отличается от обычного тем, что это препятствие в нем отсутствует. Конечно, имеют значение и другие обстоятельства, в частности число содержащихся в яйце хромосом, число выделяемых полярных телец и роль сперматозоида, заключающаяся либо в введении в яйцо центра деления, либо в вызывании там его развития. Эти взаимоотношения станут более ясными, когда будет описано поведение яиц, искусственно стимулируемых к развитию. {163}
ИСКУССТВЕННОЕ ОПЛОДОТВОРЕНИЕ
Еще в 1866 году Тихомировым было обнаружено, что неоплодотворенные яйца шелковичной бабочки при трении между двух кусков ткани или при погружении на короткое время в серную кислоту начинают развиваться (6%) и образуют зародыши. Позднее этот опыт был повторен различными путями и в целом подтвержден, но была лишь сделана оговорка, что даже и без воздействия неоплодотворенные яйца некоторых рас этой бабочки тоже могут начать развиваться. Примерно тридцатью годами позднее было обнаружено, что неоплодотворенные яйца морского ежа и одной аннелиды можно заставить развиваться по крайней мере до дробления путем прибавления к морской воде некоторых солей. Развития неоплодотворенных яиц морского ежа в плавающий зародыш не удавалось получить до 1900 года, когда Лёбу удалось вызвать это путем изменения химического состава морской воды. С тех пор было показано, что яйца можно заставить развиваться многими различными агентами. Нельзя сказать, чтобы эти опыты осуществили давнишнюю надежду, что «химическое оплодотворение» разъяснит природу нормального оплодотворения, но рассмотрение изменений, вызываемых в яйце искусственными агентами, привело к некоторым интересным рассуждениям относительно природы изменений, которые может совершать сперматозоид при проникновении в яйцо. Трудности интерпретации этой проблемы обусловлены не отсутствием фактов, а открытием весьма многих путей, по которым может начаться развитие. Установить какое-либо общее свойство для всех этих агентов оказалось невозможным. Все попытки объяснения сбивались до сих пор с толку изменениями в самом яйце. Это станет очевидным, когда будут рассмотрены некоторые из применявшихся методов.
В одном из наиболее ранних опытов яйца морского ежа помещались примерно на час в морскую воду, к которой был добавлен хлористый магний (50 см3 морской воды (50 см3 2,5-нормального раствора MgCl2), после чего они переносились обратно в морскую воду. Более совершенный метод состоял в переносе яиц на два часа в гипертоническую морскую воду, а затем в воду, содержащую немного уксуснокислого этила. При возвращении в морскую воду многие яйца развиваются нормальным образом. Подобный же результат дают также некоторые кислоты. {164} Первые указания на то, что яйцо будет развиваться, заключаются в отставании его оболочки. Это происходит также и при нормальном прободении яйца спермием. Появляется прозрачная оболочка, которая отстает от яйца и довольно скоро отделяется от поверхности его, оставляя внутри пространство, заполненное жидкостью. Раньше предполагалось, что одним из существенных факторов в начале развития яйца является образование оболочки, но с тех пор были найдены другие способы воздействия, препятствующие отставанию оболочки и тем не менее не влияющие на нормальное развитие.
Яйца морской звезды можно заставить развиваться, воздействуя на них в определенный момент морской водой, насыщенной углекислотой. Дробление яиц вызывается даже встряхиванием или воздействием масляной кислотой. Яйцо лягушки можно заставить развиваться путем накалывания его тонкой иголкой. Важно ли или нет для успеха наличие на иголке следов лимфы или крови не ясно, но некоторые опыты, поставленные для проверки значения этого момента, показывают, поводимому, что хотя в отсутствии лимфы развитие и может начаться, но фигура деления при этом не развивается, в то время как в присутствии лимфы некоторые яйца могут делиться. Из тысяч наколотых яиц в лучшем случае только одно может дать начало молодому зародышу, и очень редко случается, что какой-либо из них развивается до стадии лягушки. Яйца некоторых аннелид могут быть стимулированы к развитию хлористым калием и натрием, яйца же других — кислотами, температурой или, наконец, разбавлением морской воды.
На основании этих данных очевидно, что хотя и имеются специфические агенты, дающие наилучший результат для каждого рода яиц, но эти агенты настолько различны, что невозможно сделать общее заключение о способе их действия. Лёб придавал большое значение цитолитическому эффекту этих агентов на поверхностный слой яйца. В известном смысле эффект этот можно назвать разрушительным. Лёб предполагал, что двойное воздействие дает лучший результат, потому что второе воздействие противодействует эффекту цитолитического действия первого. Но так как во многих случаях достаточно единственного воздействия, то из этого не следует, что этот эффект является цитолитическим, а даже если это и {165} так, то природа такого изменения еще не выяснена. Необходимо выяснить, каким образом эти агенты удаляют препятствие, подавляющее изменения, которые готово претерпеть само яйцо. Несомненно, что некоторые из применявшихся агентов действовали на яйцо повреждающим образом, ибо при оставлении в этих жидкостях дольше оптимального времени, яйца безвозвратно повреждались. Начальным изменением на поверхности яйца может быть
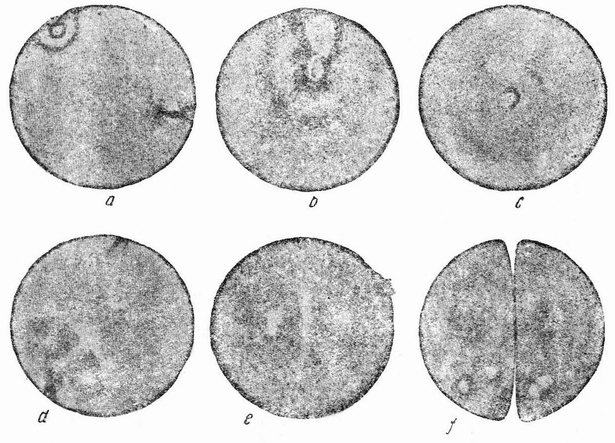 |
| Рис. 92. а, b, с — срезы через яйцо лягушки, стимулированное к развитию накалыванием чистой иголкой, видно образование звезды около места прокола; d, е, — срезы через яйца, наколотые иглой с кровью (двойное воздействие) (по Герланту). |
шаг втом же самом направлении, но термин цитолитический процесс недостаточно объясняет, что именно при этом происходит; разве только это может означать какого-то рода изменение, повреждающее какую-нибудь структуру на поверхности яйца или контролирующее его развитие.
В последние годы много внимания было уделено тем изменениям, которые могут быть видимыми в самом яйце после воздействия на него активирующими факторами. Это главным образом касается развития в протоплазме центров деления и поведения яйцевого ядра. Короче {166} говоря, было найдено, что все успешные опыты приводили к развитию в яйце звезд и что лучшими воздействиями были те, при которых вблизи ядра появлялось немного звезд, предпочтительно только две. Между звездами развивается веретено, к которому прикрепляются хромосомы, и затем может произойти деление. Надо отметить, что в нормально оплодотворенном яйце звезда появляется вблизи проникающего в яйцо ядра спермия и делится для образования полюсов веретена. Звезда, лежащая на внутреннем конце веретена второго полярного тельца, к этому моменту исчезает. Доказательства восстановления или сохранения этой звезды в яйце, искусственно стимулированном к развитию, отсутствуют, наоборот, как установлено выше, звезда появляется de novo. Более того, фигура деления не образуется путем деления одной из этих звезд, а возникает из одной или большего числа их, возникших независимо. Создается впечатление, что хромосомы посредством их прикрепляющих нитей находят эти веретена и используют их.
То, что происходит с ядром, зависит, повидимому, как от рода яиц, так и от характера воздействия. В зрелом яйце морского ежа, где образование полярного тельца закончилось, ситуация относительно проста. Гаплоидное ядро делится в нем, и каждая получающаяся клетка имеет гаплоидное же число хромосом. Клетки личинки, повидимому, тоже гаплоидны. Возможно, что случайно хромосомы могут разделиться, в то время как деления яйца не происходит. Таким образом, яйцо становится диплоидным прежде, чем происходит следующее (которое теперь является первым полным) деление и образуется диплоидный зародыш. Кажется вероятным, что те редкие случаи, в которых зародыш развивался до взрослой формы, являются как раз теми, в которых такая диплоидность имела место.
Практически тот же самый вывод применим и к яйцу лягушки. После накалывания в нем выделяются два полярных тельца, и гаплоидное яйцевое ядро отходит к центру яйца, где оно входит в соприкосновение с двумя появившимися в яйце звездами (рис. 92). Гаплоидные эмбрионы выглядят слабыми. В начале развития количество протоплазмы у них вдвое превышает то, которое соответствует числу их хромосом. Не ясно, является ли именно это первичной причиной их слабости, и неизвестно, происходит ли на более поздних стадиях регуляция между гаплоидным {167} числом хромосом и величиной клетки. Все особи, развившиеся до взрослых стадий, оказались диплоидными. Знание поведения хромосом при естественном партеногенезе делает возможным изучение вопросов наследственности. Так как в этих случаях выделяется только единственное полярное тельце и хромосомы продольно расщепляются, как и в обычном митозе, то все потомство женской линии имеет в точности одинаковое генетическое строение. Благодаря генетической идентичности этих самок, вся индивидуальная изменчивость, которую они обнаруживают, может быть отнесена за счет небольших различий во влияниях среды. Проведенные измерения действительно, обнаружили в этих случаях такое же близкое сходство, какое имеется у идентичных близнецов, причем это происходит здесь по той же самой причине, ибо в обоих случаях особи имеют идентичные гены. Наоборот, внезапные и часто большие как структурные, так и физиологические изменения, которые имеют место при окончании партеногенетического цикла и при начале половой фазы дают возможность изучить, в какой степени некоторые новые внешние влияния смогут дать новый конечный результат на том же самом генетическом фоне. Здесь опять возникает уже обсуждавшийся ранее вопрос, а именно, обусловливается ли изменение началом действия различных генов, или же продукт всех генов остается тем же, а внешняя среда заставляет эти продукты давать отличный конечный результат. Ответ и в этом случае не убедителен, но так как было показано, что изменение происходит внезапно, действуя на организм скорее как стимул, а не продолжительно в течение всего развития, то может казаться более вероятным, что на критической стадии приходят в действие различные гены.
| << | | {168} | | >> |