Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА XIV |
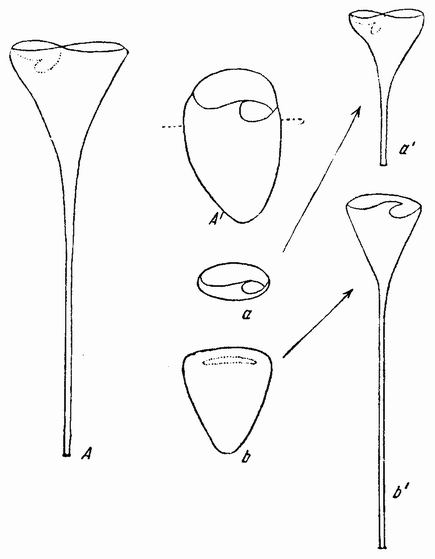 |
| Рис. 93. Stentor coeruleus в вытянутом (А) и в сократившемся (А1) состоянии. На рис. а и b показано место разреза на две части; а1 и b1 — регенерация из каждой части двух новых стенторов (по Моргану). |
не сокращается и не перестраивается сначала в новое целое меньших размеров, а новый гидрант с самого начала оказывается почти такой же величины, как и исходный. Tubularia является колониальной формой, состоящей из ветвящегося в основании ствола, образующего трубчатые стебельки, каждый из которых заканчивается головкой {170} или гидрантом, имеющим два ряда щупалец, перистом и рот (рис. 95 а). Если гидрант разрезан, то открытый конец стебелька менее чем в полчаса покрывается слоем клеток, которые выталкиваются из поверхности разреза стебелька. Покрывающие клетки являются не новыми, а представляют собой старые клетки из окружающей разрез стенки. Приблизительно через 12 часов около срезанного конца
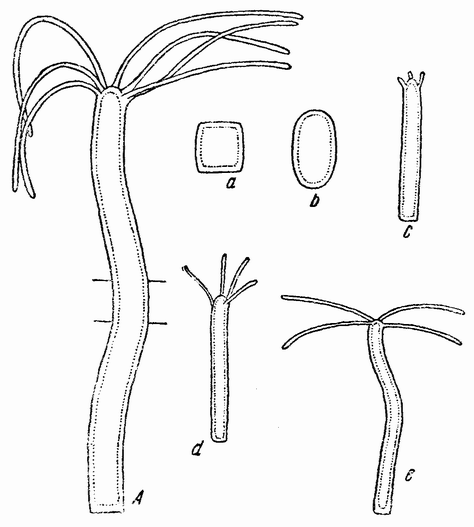 |
| Рис. 94. А — Hydra viridis; a — кусочек, вырезанный из ее середины и его развитие (b, c, d, е) (по Моргану). |
появляются два ряда пигментированных полосок (рис. 95 b), из которых в дальнейшем развиваются два ряда щупалец. Эти полоски отделяются от поверхности по всей своей длине, прикрепляясь к стебельку только своим основанием, и становятся щупальцами. Над внешним кругом щупалец образуется перистом с новым ртом на его конце. Затем новый гидрант выдвигается вперед за срезанный конец хитиновой трубки (рис. 95 с). Гистологическое {171} изучение такой части в то время, когда происходят эти изменения, показало, что новых клеток при этом либо вовсе не образуется, либо их образуется очень мало. В новые части, шупальцы, перистом и т. д., непосредственно превращаются старые клетки.
Стенки стебелька Tubularia состоят из двух слоев клеток, располагающихся с внутренней стороны кутикулы. Широкий центральный пищевод неполностью разделяется продольной перегородкой, по одной стороне ко-
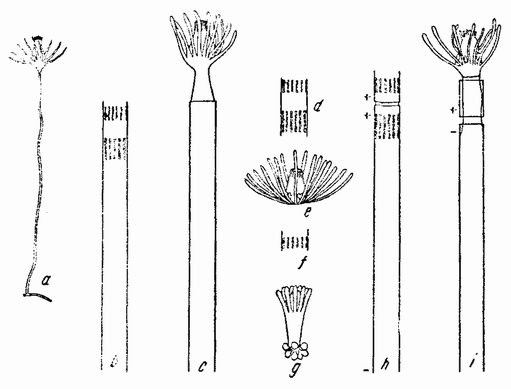 |
| Рис. 95. Регенерация у Tubularia. а — гидрант и стебелек; b — появление вблизи срезанного конца двух кругов щупалец; с — гидрант, развившийся из b, d, е — регенерация гидранта без стебелька, из короткого кусочка; f, g — регенерация только дистального конца гидранта; h — короткий кусочек, привитой на конец длинной части в обратном положении; i — регенерация из него единого гидранта (по Пиблсу). |
торой пищеварительная жидкость циркулирует вверх, а по другой вниз, проходя вокруг свободного конца перегородки как раз под гидрантом. Вскоре после удаления старого гидранта перегородка поблизости от поверхности среза разрушается, и ее клетки, содержащие зерна красного пигмента, попадают в пищеварительную жидкость. Так как при появлении щупальцевых полосок в них развивается красный пигмент, то до того, как его происхождение стало известным, предполагалось, что красный пигмент пищеварительной жидкости является формообразовательным {172} веществом. Однако этот взгляд упускает из вида тот простой факт, что во время закладки пигментированных полосок зерна пигмента циркулируют по всей длине стебля, а не собираются на его переднем конце. Только после закладки щупалец и образования головы зернышки слипаются вместе в желудке нового гидранта и извергаются изо рта, когда гидрант выдвигается вперед над старой кутикулярной стенкой. Попытка усмотреть в этом случае формообразующее вещество была явно преждевременна.
В качестве примера регенерации путем развития новых клеток может быть выбран процесс, происходящий у пресноводного кольчатого червя Lumbriculus. Если этот червь разрезан на две части, то новый хвост вырастает из заднего конца передней части, а новая голова из переднего конца задней части. Через несколько дней после разреза на срезанной поверхности появляется нарост из клеток, нг обнаруживающих диференцировки старых клеток, из которых они произошли. Они стали более округлыми и потеряли в некоторой степени их видимые морфологические черты. В деталях эти изменения заключаются в следующем: срезанный конец закрывается стягиванием стенок тела. Затем, когда стенки расслабляются, слой эктодермальных клеток наползает на конец, некоторые из этих клеток втягиваются и начинают заполнять собой внутренность. Далее эти внутренние клетки начинают диференцироваться заново. Из внутренних клеток нароста происходят мозг, соединительная ткань и брюшная цепочка формирующегося головного участка, а также мускулатура. На конце вдается внутрь трубка, образованная из поверхностной эктодермы и из которой развивается пищевод, внутренний конец которого соединяется с передним концом пищеварительного тракта и открывается в него. По мере удлинения нового нароста он обнаруживает циклические перетяжки на шесть или семь колец, или сомитов. Затем процесс заканчивается, причем замещения всего переднего конца не происходит (если было удалено больше чем семь или восемь сегментов), а восстанавливается только часть его, достаточная для образования головы.
Образование нового хвоста происходит сходным образом за исключением лишь того, что после образования нескольких первых сегментов на конце образуется растущая зона, которая прогрессивно добавляет новые сегменты {173} к образованным в начале и которая в конце концов может дать начало отрезанному числу сегментов.
Самая важная особенность регенерации новых частей у этого червя заключается в происхождении и судьбе новых клеток. Новый кожный покров происходит из клеток старого покрова, причем это относится также и к новому мозгу, нервной цепочке и даже к новой мускулатуре. Насколько известно, все эти три ткани или органа происходят из одних и тех же исходных клеток, которые раньше выполняли функции кожного покрова червя. Какая из трех возможностей реализуется, будет, повидимому, определяться локализацией или положением этих клеток в новой части. Каким из этих фактов придать большее значение, долгое время являлось загадкой для исследователей. Одно время считалось, что сходные ткани образуют сходные, но очевидно, что это верно только частично: для Tubularia это так, но у Lumbriculus это верно только для кожного покрова. Предполагалось также, что регенерирующие органы образуются таким же образом, как и в эмбрионе, т. е. из тех же зародышевых листков, однако, для мускулатуры у Lumbriculus это не верно.
Это еще раз выдвигает вопрос о роли генов в этих регенеративных изменениях. Так как все клетки содержат полный комплект генов, то, казалось бы, каждая клетка способна к образованию любой части организма, если только ее протоплазма не была уже раньше необратимо изменена в каком-нибудь определенном направлении. Исходя из этого предположения, должно казаться более вероятным, что старые клетки будут продолжать функционировать в новой части так же, как и раньше, и во многих случаях это, очевидно, соответствует действительности; но к клеткам, которые превращаются в другие ткани, это объяснение уже не будет применимо. Можно, пожалуй, сказать, что после того, как клетки теряют связь с тканями, из которых они происходят, некоторые из них могут утерять свою диференцировку (и наблюдения подтверждают это) и начинают развиваться согласно своему местоположению. В чем заключается значение «местоположения», трудно сказать, но путь для решения этого вопроса все же имеется. Новая диференцировка, по крайней мере в некоторых случаях, начинается в точке соприкосновения с той из старых тканей, которая лежит ближе всего к ней. Старая ткань действует на новую как диференциатор или организатор, {174} и мы можем, пожалуй, предположить, что это влияние является химическим. Однако проводить такое сравнение слишком далеко рискованно. Этого рода данные не помогают решить вопрос, действуют ли все гены в то время, когда в клетках происходят эти изменения, или же некоторые гены вызываются к действию новой средой, окружающей клетки в новом участке.
Как было установлено в главе I, одной из первых попыток объяснить не только как эмбриональное развитие, но и процессы регенерации является гипотеза Ру-Вейсмана, которая покоится на предположении, что при каждом дроблении яйца происходит качественное разделение веществ заключенных в хромосомах. Дочерние клетки приобретают, следовательно, различное проспективное значение. Как уже говорилось, это предположение не было в соответствии с известными уже в то время наблюдениями, что хромосомы в точности расщепляются пополам, и еще более не гармонирует с современными представлениями о генной структуре хромосом. Вейсман пытался сопоставить эту же гипотезу с фактами регенерации. Так как новая часть, развивающаяся из любого места разреза, подобна удаленной части, то он предполагал» что в каждом районе организма, после того как произошла окончательная диференцировка, остаются остаточные клетки, «назначением» которых является замещение дистальных по отношению к ним частей. Иными словами, при развитии организма используются не все клетки определенного сорта, а некоторые из них остаются в тканях, и именно из них и развивается новая часть. Эта гипотеза до некоторой степени объясняла бы также, почему сходные ткани образуют сходные с ними. В действительности же имеется большое число фактов, которые не находятся в соответствии с этим взглядом, ибо клетки, образующие новую часть, могут возникать из части тела, совсем отличной от той, которая дала начало рассматриваемому эмбриональному органу.
Прекрасный пример последней точки зрения найден в регенерации хрусталика в глазу саламандры (рис. 96). Если хрусталик удален, то новый хрусталик развивается из края радужницы. У эмбриона хрусталик образуется из внутреннего слоя покрывающей эктодермы. Следовательно, регенерировавший хрусталик развился из части тела, имеющей эмбриональную историю, совсем отличную от истории первоначального хрусталика. Более того, он {175} развился из функциональных клеток старой радужницы, а не из остаточных клеток. Можно сказать, что новый хрусталик развился из ткани, которая имеет наилучшее из возможных положений для образования такого хрусталика. С другой стороны, большинство клеток на краю радужницы содержит пигмент, тогда как клетки хрусталика прозрачны.
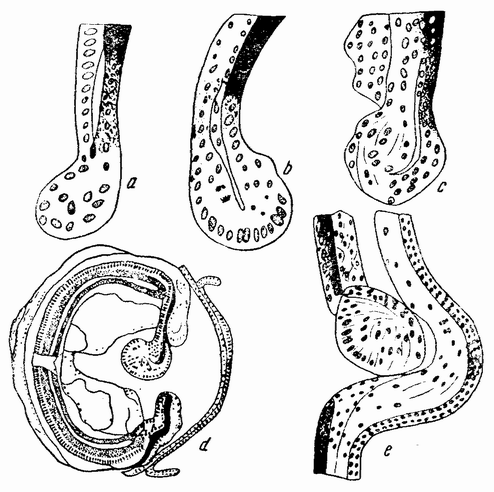 |
| Рис. 96. Регенерация глазного хрусталика у тритона после удаления старого хрусталика; a—e — стадии развития нового хрусталика, образующегося на краю радужницы; d — то же самое, видно положение нового хрусталика (по Вольфу и Фишер). |
В этом случае может казаться, что если имеются особые гены, активность которых объясняет сохранение пигментных клеток радужницы, то должны быть другие гены, которые становятся более активными, когда хрусталик теряет связь с радужницей. Но результат может быть одинаково хорошо объяснен, исходя из предположения, что в обоих случаях все гены активны и что регенерировавший орган первично детерминирован новой средой,в которой находятся клетки края радужницы. Протоплазма здесь может, следовательно, считаться отличной от той, которая {176} была раньше, вследствие чего при том же самом продукте генов конечный результат является тоже отличным. Конечно, это заключение не объясняет специфической природы реакции. А это как раз и представляет собой суть спора. Неудивительно поэтому, что, благодаря нашей неспособности установить, какие факторы участвуют в вызывании этих результатов, ряд эмбриологов прибег к сверхъестественным, т. е. метафизическим, попыткам их объяснения. Наиболее популярное из этих объяснений известно под именем витализма, причем в отдельных обсуждавшихся случаях в качестве виталистического агента призывалась энтелехия. Этот взгляд, пожалуй, даже не заслуживает критики, поскольку он основывается на метафизической концепции, но по крайней мере некоторые замечания следует все-таки сделать: во-первых, такое объяснение также трудно «понять», как и объяснить с его помощью факты; во-вторых, если с ним согласиться, то, благодаря допущению целесообразности, оно будет тормозить все дальнейшие попытки найти натуралистическое объяснение; и, в-третьих, при изучении регенерации получены некоторые факты, ставящие это учение в очень смешное положение, поскольку энтелехия выступает как благотворительный фактор.
Например, если дождевой червь разрезан по середине на две части, то на заднем участке развивается голова, состоящая из четырех или пяти сегментов, но так как область, содержащая репродуктивные органы, не регенерирует, то такое «животное» никогда не может размножиться. Если червь разрезан ближе к заднему концу, то на переднем конце задней части (рис. 97 d, е) развивается хвост, который продолжает прибавлять новые сегменты, но такая часть с двумя хвостами через некоторое время погибает, так как она не может принимать пищу.
Подобный же результат получается, когда у планарии отрезается небольшой кусочек на конце хвоста. В этом случае на переднем конце отрезанной части регенерирует другой хвост. Если у планарии отрезан конец головы, то на задней поверхности среза развивается новая голова (рис. 91 а, b, с). Двухголовая часть погибает, так как у ней не образуется рот. Получается, таким образом, что энтелехия как будто бы может делать грубые ошибки, при чем при этих условиях это происходит постоянно, что и поощряет поиски другого объяснения. {177}
У высших растений большинство регенеративных явлений настолько отлично от регенерации животных, что сравнивать их трудно. Удаление терминального конуса роста сопровождается развитием нескольких преформированных почек, которые уже присутствуют ниже по стеблю. Здесь вопрос заключается скорее в агенте, который контролирует развитие почек, причем имеются данные, доказывающие, что это может быть связано с появлением какого-то веще-
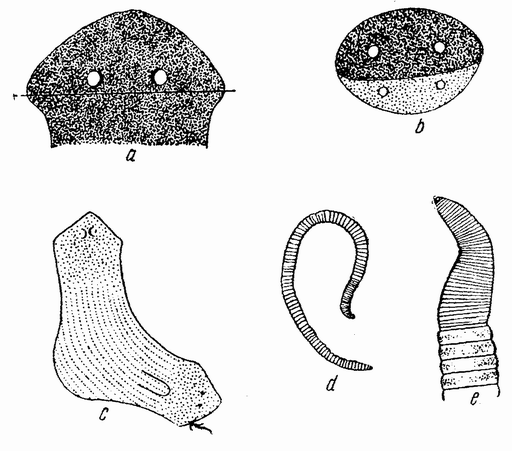 |
| Рис. 97. а — голова Planaria lugubris, видно место разреза; b — регенерация новой головы на заднем конце старой головы; с — более крупный кусочек с глоткой и двумя головами; d — хвост дождевого червя, у которого на переднем срезе регенерировал хвост; е — то же самое, увеличено (по Моргану). |
ства, вырабатывающегося в верхушке стебля. Субстанция, представляющая собой это вещество, была выделена и ее действие показано экспериментально. Следовательно, химическое вещество имеет по крайней мере некоторые свойства энтелехии.
Другие растения могут регенерировать также путем образования маленьких отростков на поверхности среза. Поверхность разреза сначала покрывается неспециализированными клетками из так называемого камбиального слоя. В одной или нескольких точках новой поверхности {178} появляется небольшая почка, которая в одних случаях, вероятно, часто происходит из единственной клетки, в других же — из нескольких клеток. Один из этих отростков может вырасти в терминальный стебель или ветку или же, если он удален и посажен в землю, то он образует корни и станет целым растением.
В двух случаях, выбранных для иллюстрации регенерации у животных путем образования новой ткани, развитие происходило из передней и задней поверхностей раз-
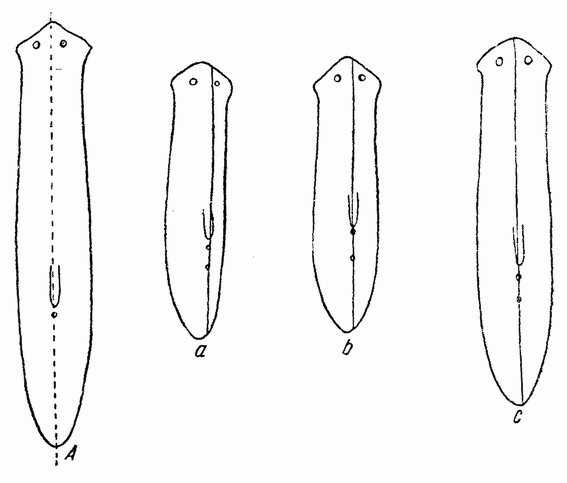 |
| Рис. 98. А — Planaria, продольно разрезанная на две части; а, b, с — регенерация целого червя из левой половины А (по Моргану). |
реза. Однако может также иметь место и латеральная регенерация, как это, например, происходит, если отрезать ногу саламандры в любом месте. Из оставшейся части конечности регенерирует ровно столько, сколько было отрезано, а отрезанный кусок погибает от недостатка питания. Если отрезана правая нога, то новая нога оказывается правой; если отрезана левая нога — новая нога будет тоже левой. Если плоский червь продольно разрезан надвое (рис. 98 А), то каждая половинка восстанавливает {179} недостающую половину. Левая половина червя (рис. 98 a) образует новую правую сторону и между старой и новой частями устанавливается срединная плоскость (рис. 98 с). То, что происходит в этом случае, подобно тому, что имеет место при разъединении первых двух бластомеров ланцетника. Если бы оба бластомера остались вместе, то один из них должен был бы образовать правую сторону тела, но после изоляции из его вещества развиваются и правая и левая стороны эмбриона, причем это достигается не путем регенерации клеток для отсутствующей половины, а путем установления в сферической бластуле новой срединной плоскости, которая была, так сказать, создана клетками правой стороны. Это же имеет место также и у Stentor, если его разрезать в продольной плоскости на две части.
Другой вопрос, который следует поставить, касается связи между скоростью роста и местом, на котором произведен разрез. Если отрезать кончики хвостов двух дождевых червей, один недалеко от конца, а другой дальше кпереди, то скорость, с которой происходит регенерация, медленнее, когда разрез расположен ближе к концу, и быстрее, когда он находится дальше кпереди. Два задних конца могут восстановить недостающие части в течение одного и того же времени, благодаря более быстрому росту от разреза, расположенного дальше кпереди. Телеологически это можно объяснить либо допущением, что чем большая необходимость в органе испытывается организмом, тем быстрее происходит рост, либо же, что энтелехия действует быстрее в передней половине, потому что там «чувствуется» большая нужда для приведения червя в порядок. Имеется, однако, биологический предел даже и для «мудрости» энтелехии, ибо если разрез сделан еще дальше кпереди, то рост заднего конца вообще не происходит.
То же самое справедливо и для конечности саламандры. Если отрезан конец одного пальца, то его восстановление будет продолжаться столько же времени, как и восстановление всей ноги, отрезанной в ее основании. Очевидно, что нужда в целой ноге больше нужды в кончике одного пальца и т. д. Оставляя в стороне метафизические толкования, факты относительно скорости регенерации в зависимости от места разреза дают материал для некоторых интересных сравнений. Клетки верхушки пальца нормальной, не {180} поврежденной саламандры или конца хвоста нормального червя прекращают размножаться, за исключением лишь размножения клеток для восстановления нормального износа частей. Можно думать поэтому, что каковы бы ни были факторы, ограничивающие максимальную возможную скорость роста в неповрежденном животном, те же самые факторы регулируют скорость роста клеток при регенерации. Неясно, однако, почему ближе к переднему концу скорость регенеративного роста должна быть больше, чем дальше от него. Этот факт требует другого объяснения.
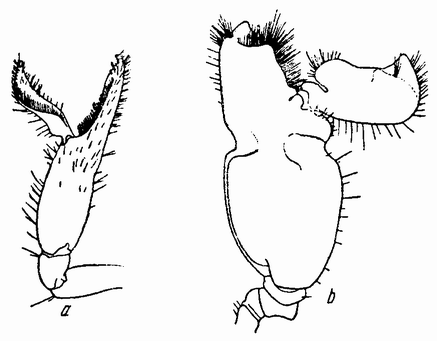 |
| Рис. 99. Alpheus. а — малая клешня; b — большая клешня (по Вильсону). |
Хорошо известно, что и при голодании кольчатые черви и планарии регенерируют. Старая, голодающая часть активно уменьшается по мере увеличения размера новой части. Но величина новой части у голодающего животного оказывается заметно меньшей, чем у питающегося. Это взаимоотношение подчеркивает тот факт, что общее количество пищи, пригодной для данной части тела, не является фактором, имеющим значение для регенерации, и сказывается только на размерах новой части. Вопрос оказывается более сложным, ибо, кроме скорости роста, еще большее значение имеет характер этого роста, а его отношение к старой части явно зависит не от количества пригодной пищи, а от других обстоятельств.
Нечто в том же роде наблюдается при компенсаторной {181} регенерации, которая может быть лучше всего иллюстрирована на регенерации большой и малой клешни ракообразного Alpheus, у которого обе клешни отличаются не только
 |
| Рис. 100. А — молодой самец краба на ранней стадии развития. В это время обе клешни более похожи на мужскую клешню; В — молодой краб, потерявший левую клешню, у которого с правой стороны регенерировала новая клешня, похожая на клешню самки (по Моргану). |
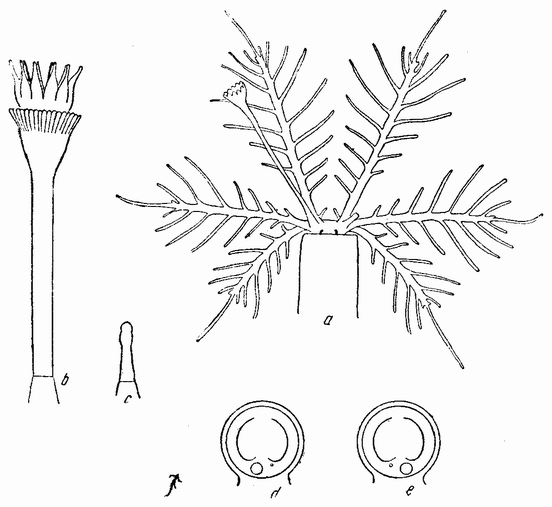 |
| Рис. 101. а — молодая стадия трубчатого червя; видно, что одна жаберная ветка изменена в крышечку; b и с — функционирующая и рудиментарная крышечка взрослого гидроида; d и е — схемы, показывающие прикрепление этой крышки (по Зеленому). |
величиной, но и формой (рис. 99). Если удалена большая клешня, то в основании ее начинает регенерировать новая. Когда животное линяет, вылезая из своей старой кутикулы, {182} то первоначальная маленькая клешня превращается в большую клешню, а регенерировавшая клешня в меньшую. Если, наоборот, удалена маленькая клешня, то новая оказывается тоже малой, а старая после линьки тоже попрежнему остается большой клешней. Повидимому, здесь имеется своего рода физиологическое равновесие между клешнями обоих сортов, природа которого, однако, неизвестна.
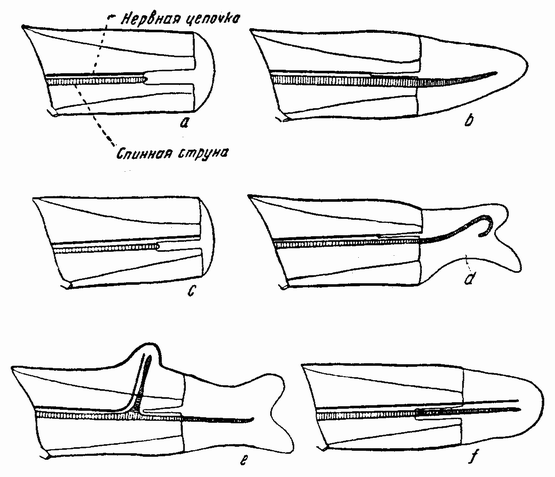 |
| Рис. 102. Хвост головастика, а — спинная струна и нервный ствол из разреза удалены; с — удалена только спинная струна (нервный ствол остался); b, d, e — хвост регенерирует, когда спинная струна достигает поверхности разреза, но не регенерирует, когда до поверхности разреза доходит только нервный ствол (с) (по Моргану и Дэвису). |
У других десятиногих раков, у которых тоже имеется различие в клешнях обеих сторон, такой перемены при регенерации не происходит. Как было показано, происхождение асимметрии у одного из них, у краба-отшельника, зависит от взаимоотношений, подобных таковым у Alpheus, но в этом случае не происходит реверсии на взрослой стадии. У молодого краба обе клешни сначала одинаковы {183} (рис. 100). Если одна из них теряется, то при следующей линьке она принимает форму малой клешни, после чего реверсия уже невозможна.
У сидячих кольчатых червей имеется пробка или крышечка (рис. 101 b), которая закрывает отверстие трубки, когда животное втягивается в нее. На противоположной стороне имеется рудиментарная крышечка (рис. 101 с). Если большая функционирующая крышечка удалена, то меньшая становится большей. Регенерировавшая крышечка остается малой. Реверсия может быть получена вторично удалением вновь регенерировавшей большей
 |
| Рис. 103. а — операция удаления из места разреза вентральной нервной струны; b — то же самое другим путем. Новая голова развивается только в том случае, если на поверхности разреза имеется нервная струна или кусочек ее (b) (по Моргану). |
крышки. Таким образом, в этом случае условия являются такими же, как и у Alpheus.
Наконец, было обнаружено, что начало развития новой части обусловливается присутствием на поверхности разреза одного из тех органов, которые принимают участие в образовании данной части. Если у головастика из поверхности среза удалена из хвоста спинная струна, то новый хвостуле регенерирует (рис. 102 с). Если, однако, конец спинной струны подходит ближе к месту разреза, то путем образования новых клеток, которые впоследствии {184} достигают обнаженной поверхности, может начать развиваться хвост (рис. 102 а—f).
Если у дождевого червя из разреза удалена брюшная нервная цепочка, то новая голова на передней поверхности разреза не развивается, а может образоваться несколько кзади, в том месте, где заканчивается старый нервный ствол (рис. 103). Регенерация ноги саламандры из отрезанного конца зависит от присутствия какого-то вещества, происходящего из кости. Если вместо того, чтобы отрезать ногу, расщепить ее с одной стороны до кости, то этим может быть вызвано развитие другой вторичной ноги. Эти особые органы — нервный ствол, спинная струна и периост — могут быть названы организаторами новой части, так как они должны присутствовать для того, чтобы могло регенерировать новое целое. Не так давно это же явление было обнаружено в некоторых опытах по прививке эмбриональных организаторов. Это послужит темой следующей главы.
| << | | {185} | | >> |