Т. Г. Морган. Развитие и наследственность.. Т. Г. Морган развитие и наследственность
 Скачать 8.83 Mb. Скачать 8.83 Mb.
|

ГЛАВА III |
 |
| Рис. 3. Зрелое яйцо лягушки. |
Веретено с хромосомами движется по направлению к полюсу яйца и принимает радиальное положение. Протоплазма на полюсе начинает выпячиваться и в образовавшийся выступ продвигается один полюс веретена (рис. 4 b). По одной хромосоме из каждой пары направляется к внешнему полюсу веретена, а их партнеры к внутреннему. Протоплазматический выступ отделяется (рис. 4 с); он содержит внешнюю группу хромосом, тогда как внутренняя остается в яйце. Выделяется первое полярное тельце.
Вокруг хромосом, оставшихся в яйце, развивается новое веретено, которое скоро принимает радиальное положение под полюсом яйца (рис. 4 d). Хромосомы, оставшиеся в яйце после выделения первого полярного тельца, в это время расщепляются, причем одна половинка каждой хромосомы {23} уходит во второе полярное тельце, а вторая половинка остается в яйце (рис. 4 е). Пока это происходит, первое полярное тельце тоже делится на две равные части. В результате возникает 4 клетки (рис. 4 f), а именно три полярных тельца и яйцо, которое произошло в результате двух клеточных делений, одно из которых отличается от всех других делений тем, что во время него хромосомы каждой пары расходятся.
Многие низшие морские животные, медузы, морские ежи, черви, моллюски и рыбы выделяют свою сперму прямо в
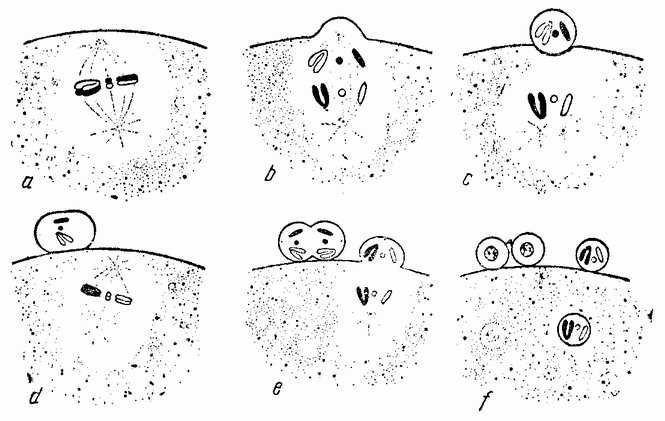 |
| Рис. 4. Деления созревания яйца; образование полярных телец. |
морскую воду, что происходит обычно одновременно с выделением яиц. Происходит потеря огромного числа сперматозоидов, но они настолько многочисленны, что все же все яйца оплодотворяются. В некоторых случаях, как например у многих саламандр, самцы откладывают сперму пачками на дно водоема, в котором в брачный период находятся оба пола. Самка ползает по этим пакетам и забирает их в клоаку. В яйцеводе пакеты растворяются, и сперма освобождается для оплодотворения яиц.
Самец лягушки обхватывает самку, влезая ей на спину, и выпускает свою сперму на яйца по мере их выхода из яйцевода. У птиц и ящериц самцы и самки совокупляются, приводя в соприкосновение отверстия клоак; сперматозоиды выбрасываются в яйцевод самки, где и происходит {24} оплодотворение. У самцов млекопитающих имеется совокупительный орган (penis), в котором проходит семяизвергательный канал. Этот орган вводится во влагалище самки, где семя испускается. Благодаря способности к движению, которой обладают сами сперматозоиды, или путем перистальтических сокращений яйцеводов сперматозоиды проходят или проводятся по яйцеводам и в их верхних отделах оплодотворяют яйца.
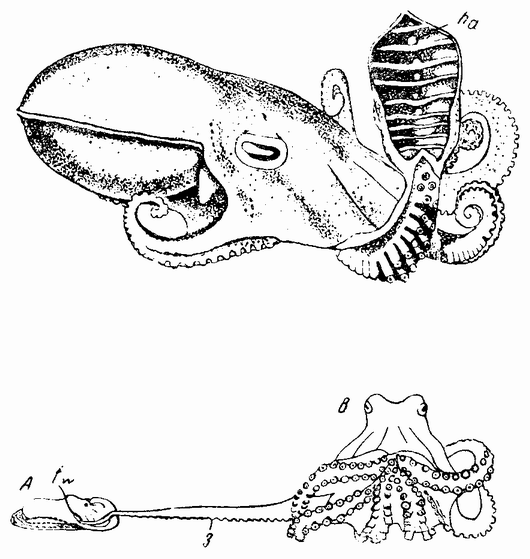 |
| Рис. 5. Самец осьминога; виден гектокотиль (ha). Копуляция (внизу): А — мелкий самец; В — крупная самка. |
Моллюски тоже обладают совокупительными органами, при помощи которых семя переносится в яйцеводы самки. У каракатиц и осьминогов перенос пакетов сперматозоидов происходит очень сложным путем (рис. 5). Особое щупальце самца переносит пакет в мантийную полость самки и сперматозоиды освобождаются из него, как только яйца выходят из яйцевода.
У дождевого червя, являющегося гермафродитом, каждая особь образует и яйца, и сперму. Во время совокупления семя одной особи попадает в особые приемники другой особи. Непосредственно перед выходом яиц вокруг тела {25} червя позади отверстия яйцеводов выделяется поясок. Этот поясок продвигается вперед, принимает в себя яйца, и в нем появляется млечная жидкость. При прохождении отверстий семяприемника сперма извергается в поясок, который продолжает затем двигаться далее к головному концу червя и, наконец, сходит с него. У коловраток самцы вводят сперму прямо в тело самок. Отдельные сперматозоиды находят себе дорогу сквозь ткани, достигая яйца в яичнике, в котором и происходит оплодотворение. У насекомых оба пола имеют в высокой степени специализированные совокупительные органы. При совокуплении семя через penis самца проходит в яйцевод самки и попадает в ее семяприемник, где и сохраняется. При прохождении каждого яйца мимо отверстия семяприемника из него выделяется небольшое количество сперматозоидов и один или большее число их проникает в каждое яйцо. Сперматозоиды могут оставаться живыми в семяприемнике самки дни, месяцы или даже годы. Так, например, пчелиная матка, которая оплодотворяется только один раз в жизни, сохраняет достаточно живой спермы для оплодотворения яиц в течение всей оставшейся жизни, которая может продолжаться несколько лет.
В яичниках самки образуются сотни, а иногда многие тысячи яиц, которые созревают либо по одному, либо все одновременно. В яичнике молодые яйца увеличиваются в числе путем обычных процессов клеточного деления и только после многих делений они претерпевают последние стадии, в течение которых число хромосом уменьшается вдвое.
У большинства животных созревание яиц приурочено к определенному времени года, когда все зрелые яйца выводятся из организма. Животные с крупными яйцами обычно откладывают их меньше, чем животные с мелкими яйцами. Некоторые птицы каждый год откладывают только два яйца, другие дюжину или больше; лягушки откладывают несколько сот яиц; рыбы и многие низшие животные, в особенности откладывающие яйца прямо в море, выделяют большие числа их. Было подсчитано, что треска производит 6 652 000 яиц, устрица — 60 000 000, моллюск Aplysia 2 000 000. Пчелиная матка производит несколько миллионов яиц, которые она по одному откладывает в ячейки сотов в течение пяти-восьми лет своей жизни.
У млекопитающих число выделяющихся из яичника яиц относительно мало. Ежегодное число новорожденных равно {26} обычно одному у рогатого скота и лошадей и доходит до двенадцати у грызунов и свиней. Некоторые мелкие млекопитающие, как например мыши и крысы, могут давать несколько пометов в год, особенно если они содержатся в тепле и хорошо питаются. У высших обезьян и человека, у которых единственный эмбрион долгое время остается у матери в матке, общее число вырабатываемых яиц относительно очень мало, несмотря даже на то, что каждое яйцо имеет микроскопические размеры. В течение жизни женщины яичники ее могут выделить около 400 яиц, которые попадают в яйцеводы, но оплодотворяются всегда только немногие из них.
Обилие вида зависит не столько от числа вырабатываемых яиц, сколько от шансов на достижение эмбрионом взрослой стадии. Поэтому животные, защищающие свою молодь, оставляя ее в теле матери или заботясь о ней после рождения, могут оставить также много потомков, как и те, которые производят много большее число яиц.
ЗРЕЛЫЕ СПЕРМАТОЗОИДЫ
Типичный сперматозоид представляет собой мелкое нитеобразное «микроскопическое животное (анималькуль)», имеющее на одном конце округлую головку и длинный хвостик (рис. 6 а—f) Это строение может быть видимо только при очень больших увеличениях микроскопа. При попадании в воду «спермий» плавает, повидимому, самым неправильным образом, с помощью колебательных движений хвостика. Неправильность его движений, несомненно, частично зависит от столкновений с другими сперматозоидами или с мелкими частицами, плавающими в воде. Иногда можно встретить описания, что сперматозоид движется по спирали в свободной среде и циклически при соприкосновении с твердым телом.
Если в воде находятся яйца, то сперматозоиды быстро собираются в большом числе вокруг них, создавая впечатление, что они притягиваются яйцами. В действительности, однако, аккумуляция сперматозоидов является в первую очередь обусловленной прилипанием их к оболочке или к слизи, окружающей яйцо. Каждый сперматозоид, соприкоснувшийся с оболочкой яйца, прилипает к ней и через очень короткое время вокруг яйца могут оказаться сотни сперматозоидов. Прямых данных о том, что плавающие вокруг яйца сперматозоиды активно поворачивают к нему, нет, {27} наоборот, есть много указаний, что сперматозоиды могут находиться поблизости от яйца и проплывать мимо него. Большинство сперматозоидов немедленно после соприкосновения с яйцом принимает вертикальное положение,
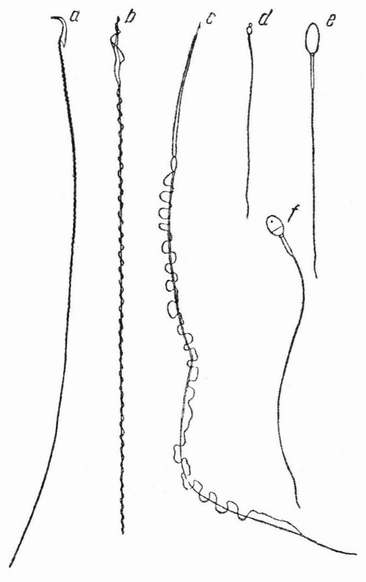 |
| Рис. 6. Форма сперматозоидов: а — мыши; b — зяблика; с — тритона; d — асцидии; e — лошади; f — человека (по Балловицу, Иенсену и Броман). |
При убивании сперматозоидов соответствующими реактивами и при специальной окраске их было найдено, что они устроены весьма сложно. Впередиголовки часто имеется особое тельце, называемое акросомой, функция которого хотя и неизвестна, но надо думать, что она имеет какое-то отношение к способности сперматозоида проникать сквозь оболочку яйца. Головка сперматозоида состоит главным образом из ядра той клетки, из которой он развился. Эта часть спермия состоит почти из одного хроматина, хромосомы оказываются уплотненными в плотную массу. Позади головки находится промежуточный участок, происходящий из протоплазмы исходной клетки. Он может содержать в себе одно или более особых телец, в частности клеточную центриоль или по крайней мере тельце, становящееся клеточным центром после проникновения {23} сперматозоида в яйцо. В длинном сплющенном хвостике можно иногда видеть осевую нить, окаймленную с каждой стороны плавникообразной перепонкой. Движение спермия обусловлено сокращениями хвостика, спиральный же путь его, возможно, зависит от некоторой асимметрии головки или хвостика. Проникновение сперматозоида в яйцо наблюдалось ча-
 |
| Рис. 7. Оплодотворение и образование полярных телец у Nereis. На четырех меньших рисунках изображено проникновение спермия. Выделение первого полярного тельца показано на левом нижнем рисунке, а выделение второго полярного тельца на двух больших рисунках справа. Последние три рисунка также изображают образование лучистости вокруг центриоли, внесенной сперматозоидом, которое является началом появления веретена первого дробления яйца (по Ф. Лилли). |
сто (рис. 7), но как ему в самом начале удается проникнуть сквозь часто очень вязкую окружающую оболочку — этого еще никогда не видели. Через несколько секунд наружная поверхность яйца обнаруживает явственный выступ протоплазмы в форме бугорка, находящегося как раз под головкой спермия. Как правило меньше чем в течение одной минуты головка проходит сквозь {29} оболочку, причем, когда половина ее уже прошла, она часто обнаруживает посредине перетяжку. Сейчас же после прохождения сквозь оболочку головка захватывается воспринимающим бугорком, который вместе с содержащейся в нем головкой спермия начинает в свою очередь втягиваться в яйцо.
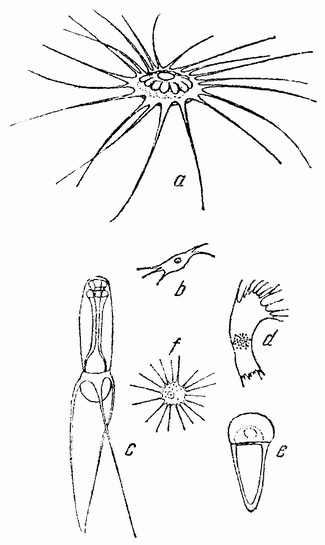 |
| Рис. 8. Необычная форма сперматозоидов: a — речного рака; b — Polyphemus; c — омара; d — Sida; e — аскариды; f — Moina (из Коршельта и Тендера). |
Хотя большинство сперматозоидов как у низших, так и у высших групп имеет нитеобразную форму, тем не менее имеется также некоторое число других типов, лишенных хвостиков. У круглого червя — лошадиной аскариды — сперматозоид имеет амебоидную форму (рис. 8 е). Оплодотворение {30} в этом случае внутреннее — семя самца переносится к яйцевод самки. У некоторых высших ракообразных сперматозоиды имеют весьма различную форму (рис. 8 a—f), причем имеются различные приспособления, с помощью которых семя приходит в соприкосновение с поверхностью яйца.
Движение ядра сперматозоида от периферии к центру яйца происходит весьма сходным образом у всех животных.
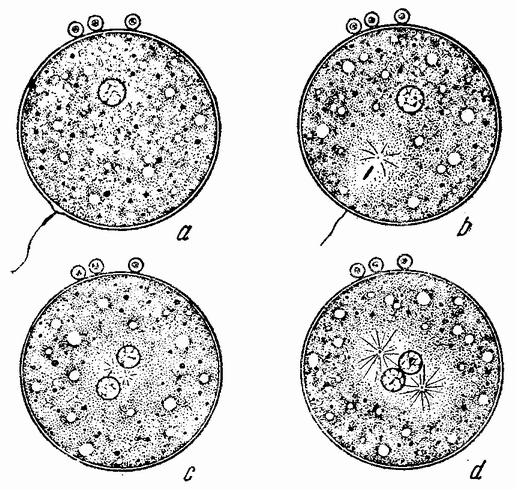 |
| Рис. 9. Оплодотворение яйца: а — сперматозоид у поверхности яйца; b — головка проникла внутрь яйца и около нее пояиилась лучистость (звезда); с — яйцевое и семенное ядро приблизились друг к другу и звезда разделилась; d — два ядра встретились, видны два полюса митотического веретена. |
Головка сперматозоида абсорбирует из яйца жидкость и медленно увеличивает свои размеры до тех пор, пока в большинстве случаев не достигнет величины яйцевого ядра (рис. 9 с—d), которое в свою очередь тоже движется от полюса яйца к центру, где оба пронуклеуса встречаются и обычно соединяются, образуя круглое ядро.
ЗАРОДЫШЕВЫЙ ПУТЬ
Происхождение половых клеток — яиц и сперматозоидов — обычно может быть прослежено, начиная с ранних стадий {31} развития эмбриона. На этом факте Вейсман построил свою знаменитую теорию изоляции половых клеток, которые рано обособляются для продолжения расы, в то время как другие клетки зародыша дифференцируются для образования сомы или тела эмбриона. Половые клетки бессмертны, соматические же клетки имеют только ограниченный срок жизни. Именно сплошным рядом половых клеток связываются современные организмы с самыми древними живыми формами, тела которых давно уже истлели. Однако,
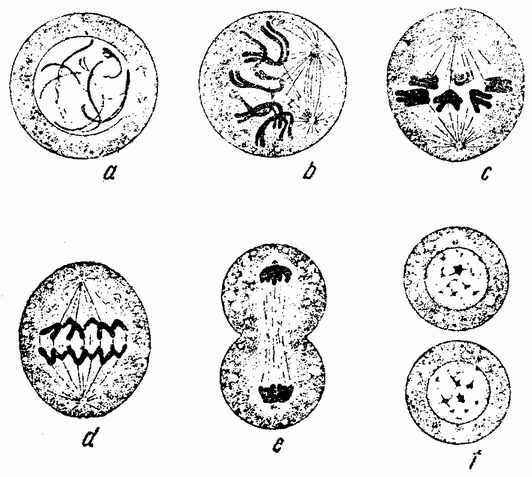 |
| Рис. 10. Стадии кариокинетического или митотического деления клетки. |
так как все наследуемые признаки расы содержатся в хромосомах всех клеток как половых, так и в клетках тела, то это различие больше не имеет того значения, которое ему придавалось Вейсманом. Тем не менее все же остается верным, что половые клетки как таковые обособляются также рано или даже еще раньше других клеток, дифференцирующихся в соматические органы эмбриона.
Вначале половые клетки не быстро увеличиваются в числе, но на более поздних стадиях развития эмбриона или у взрослого организма — время это весьма различно у разных форм — половые клетки делятся и число их увеличивается, что, особенно у самцов, может продолжаться в течение всей жизни. Эти первые деления половых {32} клеток, называемых оогониями и сперматогониями, происходят путем обычного процесса деления (рис. 10). Каждая клетка содержит в это время двойной набор хромосом, причем каждая хромосома расщепляется при каждом делении продольно на две равные половины и дочерние хромосомы расходятся в разные клетки. После того, как половые клетки претерпели ряд простых делений, они проходят два особых деления, в одном из которых число хромосом уменьшается на половину. Эти деления называются делениями созревания или, более специально, мейозом, в противоположность митозу или обычному процессу клеточного деления. Оба эти деления созревания яйцевой клетки происходят после того, как она накопила большее или меньшее количество желточного вещества. Эти два деления были уже описаны: они связаны с выделением двух полярных телец. Деления созревания в семенных клетках в основных своих чертах, поскольку это касается хромосом, происходят таким же образом, но из каждой материнской клетки происходит четыре функционирующих клетки вместо одной, как в случае яйца. Эти деления будут сейчас описаны.
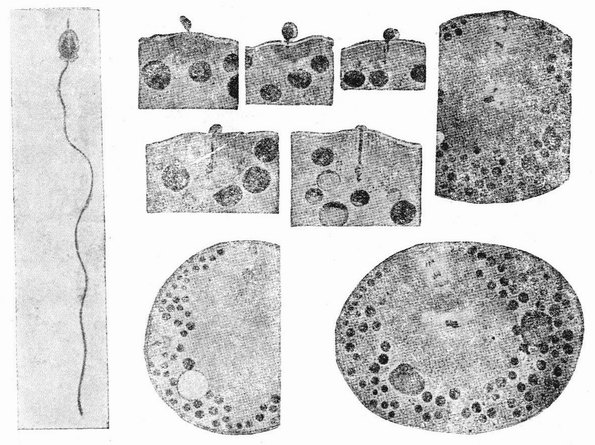
Непосредственно перед делением созревания семенных клеток хромосомы в них вновь принимают вид тонких нитей (рис. 11, a), занимающих все крупное ядро. Некоторые из хромосом имеют вид петель, другие же палочкообразны. Они располагаются так, что вдоль всей длины лежат бок о бок (рис. 11, b—с) и кажутся слившимися, но из генетических фактов известно, что в действительности они не сливаются. Является вероятным, что именно на этой стадии в яйцах или в спермиях, или в тех и в других происходит обмен между в точности эквивалентными отрезками или участками хромосом. Этот процесс называется кроссинговером (рис. 12, а—b) и в результате его возникает перекомбинация генов в гомологичных хромосомах. Далее происходит укорочение нитей и исчезновение ядерной стенки. В протоплазме появляется веретено и хромосомы прикрепляются к его нитям (рис. 11). Хромосомы выглядят еще так, как будто имеется только половина от свойственного данному виду количества, и в это время часто можно видеть, что они продольно расщеплены; иногда можно наблюдать и вторичную щель, образующую так называемые тетрады.
Затем происходит деление клетки (рис. 11, е—f—g), при котором соединившиеся пары разделяются, причем оказывается, что в некоторых случаях разделение может {33} произойти по вторичной щели или в одних хромосомах так, а в других иначе. Далее следует короткая стадия покоя и каждая клетка делится снова (рис. 11, h—i). В результате этих двух делений образуется четыре клетки, каждая из которых содержит один набор хромосом, т. е. по одной хромосоме каждого сорта. После этого следует сложный
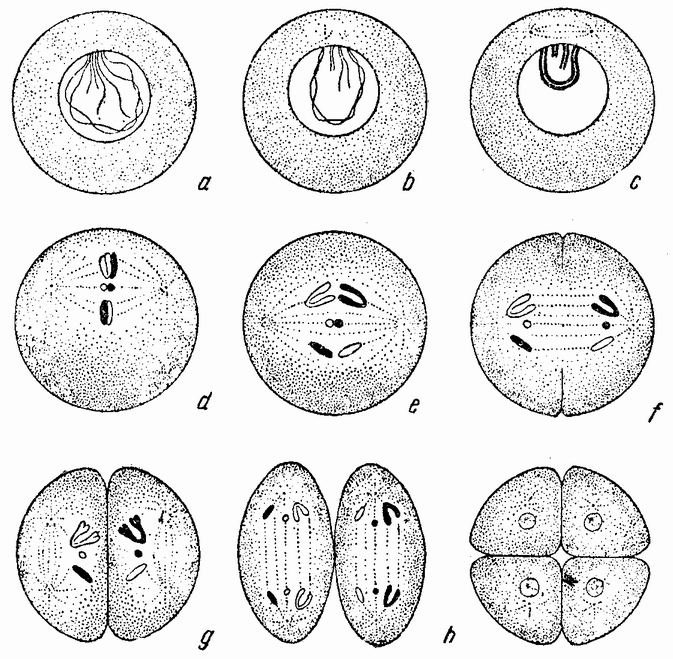 |
| Рис. 11. Два деления созревания (мейоз) семенной клетки, образующих четыре сперматозоида. |
процесс диференцировки, превращающий каждую клетку в функционирующий сперматозоид.
Является очевидным, что в отношении редукции числа хромосом деление созревания яиц и спермиев в основном сходно. Было показано как наблюдениями, так и путем дедукции из генетических фактов, что разделение конъюгирующих членов каждой пары хромосом происходит случайно. Перераспределение унаследованных материнских {34} и отцовских хромосом при делении созревания имеет место и в яйцевых, и в семенных клетках.
Значение этих фактов для теории генетики лучше всего видно в тех случаях, когда особь является гибридом, то есть когда она получает от матери один гаплоидный набор хромосом, а от отца другой, причем отличный от первого. Когда яйцо близко к созреванию, хромосомы обоих наборов соединяются в пары и если (описанная выше) редукция числа хромосом происходит случайно, то конеч-
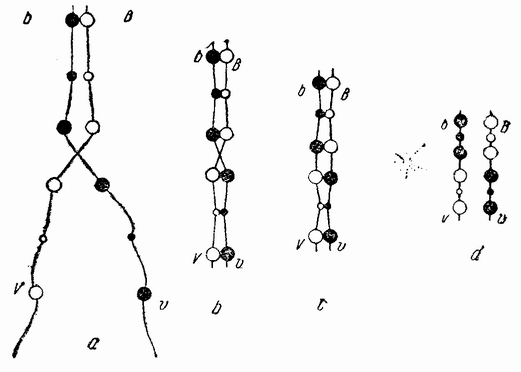 |
| Рис. 12. Схема, показывающая, каким образом две хромосомы, соединяясь вместе, могут перекрывать друг друга и обмениваться сегментами (кроссинговер). |
ный результат будет таков, что остающееся в яйце половинное число хромосом должно содержать в себе некоторые материнские и некоторые отцовские хромосомы, так как при случайном распределении редко все хромосомы будут оказываться только материнскими или только отцовскими. Таким образом, в каждом поколении зрелые яйца гибридной особи будут содержать все возможные комбинации хромосом, полученных от матери и отца, по одной каждого сорта. В потомстве, взятом в целом, будут все возможные комбинации прародительских хромосом. Этот процесс распределения хромосом и их соединения при оплодотворении представляет собой механизм двух менделевских законов наследственности.
| << | | {35} | | >> |