В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
Экология. Большинство коловраток живет в пресных водах, причем они делятся на донные и планктонные формы. Донные коловратки большей частью ползают или плавают у дна, но среди них имеется и целый ряд сидящих видов, последние прикрепляются выделением цементных желез ноги к субстрату, причем нога изменяется в своего рода стебелек. Большинство сидячих коловраток выделяет вокруг себя защитную трубку из прозрачного студенистого вещества или склеенных вместе в правильные комочки испражнений коловратки, которыми трубки обложены как микроскопическими кирпичами. У типичных планктонных коловраток на теле вырастают различного рода придатки, увеличивающие поверхность животного. У некоторых это длинные неподвижные шипы панциря (Notholca], у других — подвижно сочлененные с телом гребные щетинки (Polyarthra) или это особые, снабженные мышцами и усаженные щетинками выросты самого тела (Pedalion). Пресноводные коловратки играют немаловажную роль как пища различных, более крупных пресноводных организмов. Морских коловраток значительно меньше. Небольшое количество коловраток приспособилось к жизни на суше, главным образом во влажном мху, среди лишайников и т. п. Некоторые из них (Callldina) охотно поселяются в особых мешочках на слоевище печеночных мхов, в которых долго удерживается вода. Наземные, а также некоторые водные коловратки способны выдерживать полное высыхание, не погибая. При этом они впадают в анабиотическое состояние (мнимая смерть), но после осторожного смачивания оживают. Оживание коловраток было впервые открыто Левенгуком в 1701 г. и с тех пор многократно исследовалось. При применении ряда предосторожностей смоченные водой коловратки оживают после многолетнего высушивания. Мало того, высушенные коловратки способны в течение 4 ч выдерживать понижение температуры до —270°С (в жидком гелии) и в течение 5 мин повышение температуры до +100°С. Филогения коловраток. О происхождении коловраток существовали различные точки зрения. Долгое время их считали неотеническими личинками каких-то кольчатых червей на основании сходства Rotatoria с личинкой Polychaeta трохофорой (см. рис. 225). В последние годы установилось мнение, что сходство коловраток с трохофорой следует считать результатом конвергенции, или совпадения, связанного с одинаковым образом жизни, путь же эволюции группы Rotatoria идет от прямокишечных турбеллярий. Защитники этого мнения считают наиболее примитивными не плавающих, а ползающих коловраток и исходным типом не хорошо дифференцированные венчики, а 240 сплошную брюшную площадку ресниц, окружающую рот. Эта площадка— остаток общего мерцательного покрова Rhabdocoela. О близости к турбелляриям свидетельствует примитивное строение протонефридиев коловраток и брюшное положение рта. Из первичнополостных червей к кл. Rotatoria ближе всего стоит кл. Gastrotricha. Их сближает отсутствие кожко-мускульного мешка, наличие протонефридиев, некоторое сходство в строении мерцательного аппарата. Дополнения к типу Nemathelminthes КЛАСС ПРИАПУЛИДЫ (PRIAPULIDA) Небольшая группа морских червей (5 родов с 7 видами), принадлежность которых к типу Nemathelminthes еще не вполне обоснована. Донные животные, обитающие в иле и песке. 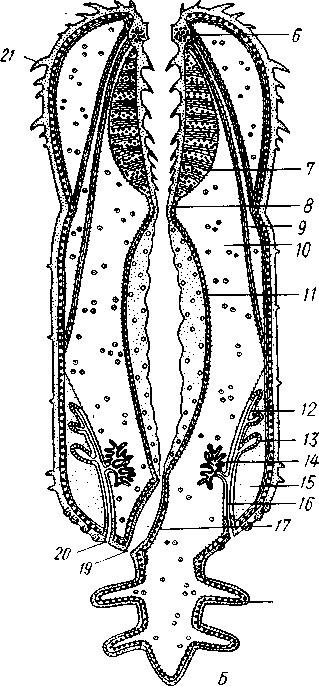 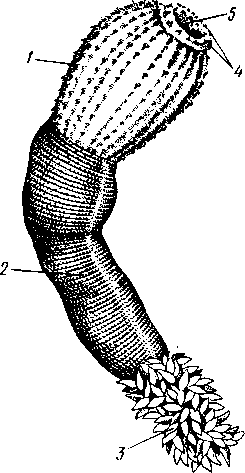 18 Рис. 204. Строение приапулиды Priapulus caudatus. A — внешний вид животного (по Шипли); Б — схема организации (по Ланду): / — хобот, 2 — туловище, 3 — хвостовые жабры, 4 — околоротовые зубчики, 5 — рот, 6 — окологлоточное нервное кольцо, 7 — глотка, S — пищевод, 9 — ретракторы хобота, 10 — полость тела, 11 — средняя кишка, 12 — половой проток, 13 — половые мешочки (части половой железы), 14 — протонефридии, 15 — брыжейка, 16 —-мочеполовой проток, 17 — задняя кишка, 18 — хвостовые жабры, 19 — анальное отверстие, 20 — мочеполовая пора, 21 — шипы хоботка Строение. Тело цилиндрическое, червеобразное, у некоторых видов снабженное 1 или 2 хвостовыми жаберными придатками (рис. 204, Л). Передняя треть животного может вворачиваться внутрь при помощи особых мускулов-ретракторов. Вворачиваемый участок тела усажен продольными рядами сосочков и шипов. Стенки тела состоят из кутикулы, однослойного эпителия и мощного слоя мускулатуры — кольцевой и продольной. Под мышцами имеется пе-ритонеальный эпителий, выстилающий обширную полость тела ', которая продолжается и в хвостовые придатки. Через все тело проходит прямой кишечник (рис. 204, Б),начинающийся на переднем полюсе ртом и заканчивающийся порошицей на заднем (у основания хвостовых придатков, если таковые имеются). Кровеносная система отсутствует. Дыхание происходит либо всей поверхностью тела, либо через хвостовые придатки, играющие роль жабр, состоящих из полого ствола, снабженного короткими веточками. Нервная система слабо развита и состоит из окологлоточного нервного кольца и брюшного нервного ствола. Выделительные и половые органы соединены у приапулид в общую мочеполовую систему (рис. 204, Б).Приапулиды раздельнополы. Половые железы образуют два продольных компактных тела, соединенных со стенками тела при помощи брыжейки. Каждая половая железа состоит из значительного числа переплетающихся между собой половых мешочков, на одном конце слепо замкнутых, а другим открывающихся в общий мочеполовой проток. Оба протока сообщаются с внешней средой при помощи пары отверстий на заднем конце тела. Вдоль мочеполового протока на нем сидит несколько кустиков протонефридиев, причем каждый кустик снабжен несколькими тысячами жгутиконосных клеток, или соленоцитов. Протонефридии открываются в мочеполовой проток. Развитие. Дробление яйца приапулид, как это недавно показано Л. Н. Жинкиным, билатеральное, т. е. напоминает дробление у первич-нополостных червей. Из яиц развиваются личинки, ведущие такой же донный образ жизни, как и взрослые черви. У Priapulus caudatus личинка устроена очень просто и состоит из эктодермы, внутри которой находится недифференцированная клеточная масса. У Halicryptus spinulosus личинка организована как взрослое животное (см. рис. 198, В); отличие лишь в том, что туловище личинки (кроме вворачиваемой передней части) одето хитиновым панцирем из двух створок: спинной и брюшной, отчего тело сильно сплющено в дорзовентральном направлении. Превращение во взрослую форму сопровождается линьками и может продолжаться до двух лет. Экология. Приапулиды живут в прибрежной зоне, делая норки в мягком грунте. Один вид обитает в придонном слое нитчатых водорослей. Размеры приапулид невелики — от 0,2 до 10 см. Филогения. Раньше класс Priapulida сближался с кольчатыми червями (кл. Echiurida и кл. Sipunculida; см. с. 281) ввиду сходства внешнего вида и образа жизни, а также потому, что у обеих групп передний конец тела может вворачиваться при помощи ретракторов, а полость тела и нервная система имеют сходное строение. В последнее время указывают на возможность сближения класса Priapulida через киноринх со стволом нечленистых червей, т. е. с Nemat-helminthes. В самом деле, личинка приапулид похожа на взрослых кино- ' Наличие специальной выстилки сближает полость тела приапулид с вторичной полостью тела — целомом кольчатых червей (см. с. 255). 242 ринх (см. рис. 198) как образом жизни, так и строением: кутикуляриза-ция покровов, наличие переднего участка тела, вооруженного несколькими венчиками шипов. И, напротив, нет никаких указаний на образование у представителей кл. Priapulida планктонных личинок трохофорного типа, столь характерных для кольчецов. В организации взрослого животного приапулид сближает с киноринхами наличие протонефридиев. Серьезным отличием от этой группы является характер полости тела, первичной у Kinorhyncha и напоминающей вторичную у приапулид. Может быть, следует считать класс Priapulida слепо заканчивающейся верхней ветвью группы нечленистых низших червей, приобретших целом независимо от типа Annelida (с. 250). Филогения типа Nemathelminth.es Долгое время происхождение классов типа Nemathelminthes оставалось неясным, и мнения авторов по этому вопросу сильно расходились. Лишь в последнее время, после более углубленного изучения групп, относящихся к этому типу, стало ясно, что все они своими филогенетическими корнями связаны с ресничными червями. В самом деле, признаки организации, общие с ресничными червями, рассеяны в разных классах типа Nemathelminthes. Так, у примитивных коловраток и брюхоресничных червей сохранились еще участки тела, покрытые ресничками. В этих классах органы выделения представлены типичными протонефридиями. Нервная система нематод (особенно у примитивных свободноживущих морских форм) и гастротрих не противоречит основной схеме строения таковой ресничных червей. Глотка пер-вичнополостных червей, в ее более примитивных формах (классы Gast-rotricha, Nematoda и Kinorhyncha), сходна с массивной глоткой прямокишечных турбеллярий. Положение рта на переднем конце тела, характерное для первичнополостных, встречается в пределах отряда Rhabdocoela. В строении половой системы, если не считать раздельнополого состояния у многих представителей типа Nemathelminthes, нет принципиальных отличий от примитивных форм из класса Turbellaria, a у коловраток женский половой аппарат снабжен желточником. Имеются, следовательно, все основания считать, что предками типа Nemathelminthes были какие-то ресничные черви. Значительными отличиями от ресничных червей в организации первичнополостных являются наличие первичной полости тела и задней кишки с анальным отверстием. Однако эти признаки являются результатом прогрессивной эволюции и не препятствуют принятию вышеизложенной теории происхождения типа Nemathelminthes. Интересно, что у некоторых прямокишечных турбеллярий лакуны в паренхиме настолько велики, что в них могут свободно плавать паразитические инфузории. Увеличение и слияние лакун могло привести к появлению обширной первичной полости тела. В строении отдельных классов типа Nemathelminthes имеются, на первый взгляд, значительные отличия от турбеллярий. Для первичнополостных червей характерна сильная кутикуляризация и более или менее полное исчезновение ресничного покрова, а также установление постоянства клеточного состава (коловратки и отчасти нематоды). У нематод отсутствует кольцевая мускулатура, в нервной системе образовалось окологлоточное кольцо, а протонефридии заменены выделительными каналами кожного происхождения. У коловраток выработался коловраща-тельный аппарат; очень сильным изменениям подверглась нервная система, глотка превратилась в жевательную, произошло распадение кож- 243 но-мускульного мешка на отдельные мышцы и т. д. У брюхоресничных и кииоринх кожно-мускульный мешок также дал начало специализированным мышцам и т. д. Все эти отличия, однако, касаются главным образом отдельных органов и тканей и мало затрагивают основной план строения. ТИП СКРЕБНИ (ACANTHOCEPHALES) Скребни — паразитические черви. Тип насчитывает около 500 видов и характеризуется следующими признаками: 1. Исключительно паразитические черви, в половозрелом состоянии обитающие в кишечнике позвоночных животных. 2. Передний конец тела преобразован в специализированный орган прикрепления в виде вворачивающегося хоботка, вооруженного рядами кутикулярных крючьев. 3. Покровы представлены тонкой кутикулой и гиподермой, пронизанной системой лакун. Полость тела первичная. 4. Пищеварительная, кровеносная и дыхательная системы отсутствуют. 5. Скребни раздельнополы. Выводные протоки половой системы нередко соединяются с протоками выделительной системы (устроенной по типу протонефридиев) в общий уро-генитальный канал. 6. Развитие с метаморфозом. Жизненный цикл проходит со сменой животных хозяев. В тип скребней входит только один класс с тем же названием. КЛАСС СКРЕБНИ (ACANTHOCEPHALA) Строение и физиология. Тело скребней имеет более или менее веретеновидную форму (рис. 205) и подразделяется на собственно тело и хоботок, полый внутри и способный вворачиваться, как палец перчатки (рис. 206). Хоботок усажен несколькими рядами загнутых назад кутикулярных крючьев и служит для прикрепления паразита к стенкам кишки хозяина. При помощи мышц-ретракторов хоботок вворачивается 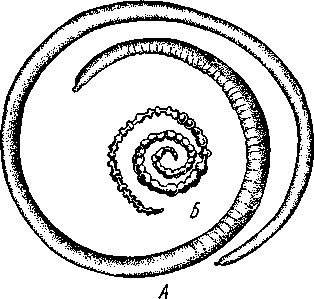 внутрь тела в особое влагалище — полый мускульный мешок, слепо замкнутый на заднем конце. Само влагалище, в свою очередь, прикрепляется к стенкам тела червя при помощи парных мышц. Тело одето тонким покровом, под которым находится гиподерма, представляющая собой синцитий, т. е. результат слияния клеток эмбриональной эктодермы. Гиподерма пронизана системой каналов, или лакун. В толще гиподермы залегают немногочисленные и довольно крупные ядра. Здесь же откладываются и запасы гликогена, который является для скребней, как и для других эндопаразитов j-, , ,. Ha границе между хоботком и соб- ственно телом скребней гиподерма об- Рис. 205. Скребни. А — гигантский (с. 173), основным источником энер-скребень Macracanthorhynchus hiru-dinaceus из свиньи; Б — скребень чет-ковидный Moniliformis moniliformis из ежа (по Бремзеру) 244 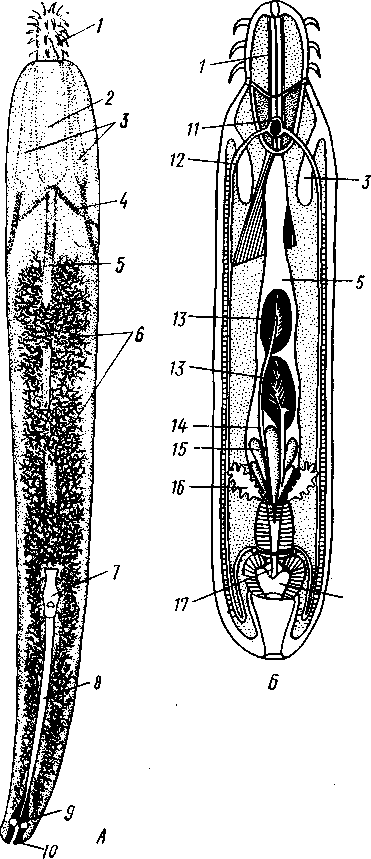 разует парные выпячивания, так называемые лемниски, которые в виде двух лентовидных тяжей лежат по обе стороны влагалища хоботка. Функция лемнисков до сих пор остается неразгаданной. Предполагается, что они принимают участие в процессах вворачивания и выворачивания хоботка. Кожно-мускульный мешок состоит из двух слоев: наружного — из кольцевых и внутреннего — из продольных мышечных волокон. Кнутри от мышц помещается обширная полость тела (рис. 207,5). Пищеварительная система отсутствует в связи с паразитизмом, и питание совершается через покровы тела. По-видимому, скребни, подобно ленточным червям, воспринимают продукты пищеварения хозяина из содержимого его кишечника. Возможно, что важную роль играют при этом бесчисленные тончайшие канальцы, открывающиеся наружу микропорами, которые пронизывают поверхностный слой покровов скребней. 18 Нервная система очень проста. Она состоит из непарного мозгового ганглия, залегающего внутри влагалища хоботка, ближе к его заднему концу. От ганглия отходят тонкие нервные веточки вперед, к хоботку, а кроме того, два более толстых боковых нервных ствола, выходящих из влагалища хоботка, пересекающих полость тела и затем направляющихся вдоль стенок тела назад. Из органов чувств описаны лишь два небольших чувствительных сосочка — у основания хоботка и на его вершине, а также возле полового отверстия. Рис. 206. Организация скребней. А — схема Acanthocephaius lucii (по Стрелкову); Б — схе* ма строения самца скребня (по Кестнеру): / — хоботок, 2 — влагалище хоботка. 3 — лемниски, 4 — мышцы, прикрепляющие влагалище к стенкам тела. 5 — лигамент, 6 — яйца и лицевые комки, 7 — маточный колокол, 8 — матка, 9 — влагалище, 10 — половое отверстие, 11 — мозговой ганглий, 12 — нервные стволы, 13 — семенники, 14 — семяпроводы, 15 — цементные железы, 16 — протонефридии, 17 — совокупительный орган, IS — совокупительная сумка V 245 Органы выделения найдены лишь у немногих видов. Например, у гигантского скребня (Macracanthorhynchus hiru-dinaceus) они имеют характер несколько видоизмененных протонефридиев. Это два древовидно разветвленных пучка, состоящих из множества тонких веточек, каждая из которых заканчивается колбовид-ной клеткой с мерцательным ,пламенем, обращенным в про- свет канальца (рис. 207,Б). Канальцы впадают в главные собирательные каналы протонефридиев, открывающиеся в выводные протоки половой системы. Таким образом, у скребней выделительная и половая системы соединены. Половая система. Скребни раздельнополы. Половые органы парные, и развитие их связано с особым соединительным тяжем, или лигамен-том, который тянется от заднего конца хоботкового влагалища через полость тела к заднему концу животного. Самец (см. рис. 206, Б) имеет два овальных семенника с семяпроводами. Оба семяпровода сливаются в семяизвергательный канал, в который вливается содержимое нескольких (чаще 6) цементных одноклеточных желез. Секрет желез служит для замыкания женского полового отверстия после совершившейся копуляции. Конец семяизвергательного канала окружен мощной мускулатурой и представляет совокупительный орган. Последний вдается в особый мешочек — совокупительную сумку, которая открывается наружу на заднем конце тела. Сумка может выворачиваться наружу и при копуляции охватывает задний конец тела самки. Женская половая система у молодых особей состоит из двух яичников, залегающих внутри лигамента. С возрастом яичники распадаются на значительное количество яйцевых комков, которые сначала поступают внутрь лигамента, а затем через разрыв стенки последнего в полость тела. Каждый яйцевой комок состоит из нескольких десятков молодых яйцевых клеток. По мере созревания яиц они оплодотворяются еще в яйцевых комках, а затем отделяются от ком-  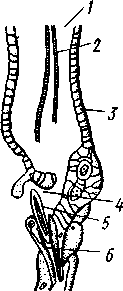 10 Рис. 207. Анатомия и гистология скребней. А — женский половой проток в продольном разрезе (по Кайзеру); Б — поперечный разрез скребня в задней трети тела (по Стрелкову); В— строение протонефридия (по Петроченко): |