В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
Развитие. Дробление яйца полное. Сначала двумя последовательными меридиональными делениями яйцо рассекается на 4 приблизительно одинаковых бластомера: А, В, С и D, причем В отвечает будущей брюшной, a D — будущей спинной стороне зародыша. Затем эти 4 клетки делятся наклонной бороздой на 4 мелкие верхние (анимальные) и 4 нижние крупные (вегетативные) клетки. Получается, как говорят, квартет микромеров и квартет макромеров (рис. 224). Микромеры, смотря по их происхождению от того или иного макромера, обозначают как la, \b, lc 261 и Id (рис. 224),макромеры—\А — ID. Дальнейшее дробление состоит в последовательном отделении от макромеров по направлению к ани-мальному полюсу вслед за первым еще 2-го, 3-го и 4-го квартетов микромеров, при отделении каждого нового квартета клетки ранее образовавшихся квартетов тоже делятся.  Рис. 223. Размножение полихет. А — бесполое размножение по-лихеты Autolytus (по Грассе); 5 и В — получающиеся посредством концевого почкования половые особи (5 — самка и В — самец) тех же червей (по Свешникову) Важная особенность отделения микромеров заключается в изменении направления веретена деления: анимальный конец его отклоняется то по ходу часовой стрелки, то против нее, если наблюдение вести с ани-мального полюса яйца. Если при образовании 1-го квартета ядерные веретена в бластомерах наклонены в одну сторону, то при отделении 2-го квартета все они оказываются отклоненными в другую, и т. д. Вследствие этого верхние продукты деления ложатся каждый раз не прямо над нижними, а несколько чередуясь с ними, как бы в шахматном порядке. Например, микромер \а лежит не целиком над макромером 1Л, но отчасти налегает и на соседний макромер 1В. Такое дробление называется спиральным, так как если во время деления микромера мысленно продолжить ось ядерного веретена по сферической поверхности яйца, то получится спиральная линия. 262 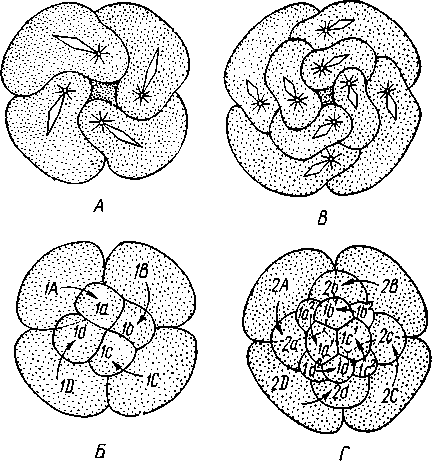 Рис. 224. Схема спирального дробления. Вид с верхнего полюса зародыша. А — переход от четырех- к восьмиклеточной стадии; Б — стадия восьми клеток; В — переход к стадии 16 клеток; Г — зародыш на стадии 16 клеток (из Зивинга) Уже на ранних стадиях дальнейшее развитие каждого эластомера точно определено. В общем, первые 3 квартета микромеров идут на образование эктодермы личинки, один из микромеров 4-го квартета — потомок клетки D, обозначаемый 4d, производит целоми-ческую мезодерму, а остальные микромеры 4-го квартета и все макромеры дают энтодерму. Ввиду такого раннего определения дальнейшего развития бластомеров дробление кольчецов называется детерминативным. В результате дробления получается шаровидная бластула, из которой посредством погружения внутрь бла-стоцеля группы крупных эн-тодермальных клеток происходит гаструла. Бласто-пор гаструлы помещается на ее вегетативном полюсе. Затем бластопор вытягивается по одной из сторон гаструлы (будущей брюшной) к ее экватору, а сзади замыкается. Его передняя часть превращается в ротовое отверстие. На нижнем полюсе зародыша образует- ся порошица. Постепенно зародыш превращается в характерную для полихет личинку — трохофору. Трохофора (рис. 225, А)—типичная планктонная личинка, плавающая при помощи ресничек. Тело ее имеет более или менее шаровидную или эллиптическую форму. На переднем (анимальном) полюсе личинки развивается чувствительный теменной султан длинных ресниц, сидящих на группе эктодермальных клеток — теменной пластинке. По экватору личинки впереди рта расположен характерный предротовой венчик ресничек— прототрох. Иногда позади рта развит менее мощный послеро-товой венчик. Кишечник начинается ртом посредине брюшной стороны личинки и заканчивается порошицей на ее заднем полюсе и состоит из трех отделов, причем передняя и задняя кишка образуются впячиванием эктодермы, а средняя кишка формируется из энтодермы. Между кишечником и стенкой тела находится первичная полость тела, пересекаемая тонкими мышечными волокнами. По бокам кишечника лежит пара маленьких протонефридиев. Мезодермальные органы личинки (главным образом мускульные волокна) развиваются из нескольких клеток, лежащих у краев бластопора и называемых мезенхимой. Другой мезодермальный зачаток представлен двумя крупными мезодермальными клетками — первичными мезоб-ластами (иногда их называют телобластами)—потомками бластомера 4d, лежащими по бокам от кишечника. После некоторого периода планктонной жизни трохофоры начинается ее метаморфоз. Заднее (вегетативное) полушарие личинки значитель- 263 но вырастает в длину и подразделяется сразу на несколько (3, 7, 9—13) сегментов. На сегментах развиваются параподии и щетинки или появляются ресничные пояски. К этому времени обе первичные мезодермальные клетки, усиленно размножаясь, дают два лежащих по бокам от кишечника клеточных тяжа — мезодермальные полоски (рис. 225, Б).Вско- 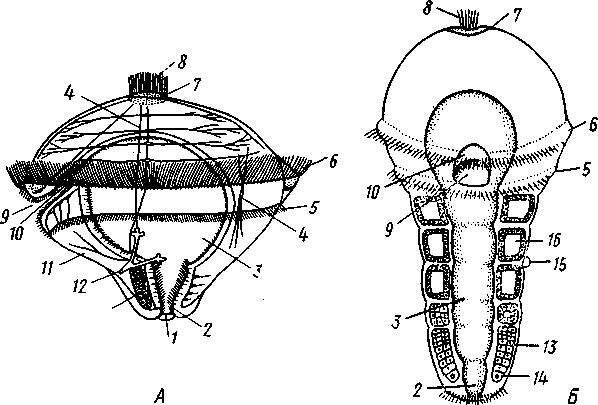 13 Рис. 225. Развитие Poltjgordius. A — трохофора, вид сбоку (по Гатчеку); Б — метаморфоз трохофоры (по Вурмбаху): 1 —анус, 2 — задняя кишка, 3 — средняя кишка, 4 — мышцы, 5 — послеротовой и 6 — предротовой венчик ресничек (нрототрох), 7 — теменная пластинка, 8 — теменной султан, 9 — рот, 10—передняя кишка, 11— нижнее полушарие трохофоры, 12—протонефридий, 13 —мезодермальная полоска, 14 — мезобласт, 15 — диссепи- мент, 16 — целом ре под влиянием наружной сегментации мезодермальные полоски расчленяются на парные группы клеток, так что в каждом сегменте оказывается своя пара мезодермальных клеточных зачатков. Последние сначала компактны, затем в них появляется полость — зачаток вторичной полости тела, а ограничивающая ее клеточная стенка есть стенка цело-мического мешка. В каждом сегменте, таким образом, развивается пара целомических мешков. В процессе метаморфоза часть клеток теменной пластинки трохофоры погружается под покровы и образует головной мозг. На брюшной стороне в виде парного валика эктодермы закладываются брюшные нервные стволы. В дальнейшем они вступают в связь с головным мозгом при помощи окологлоточных коннективов. Из эктодермы развиваются и органы чувств — глаза, пальпы. Так, из несегментированной, первичнополостной трохофоры формируется следующая личиночная стадия — метатрохофора (рис. 226), характеризующаяся сегментацией и метамерным целомом. Тело сформированной метатрохофоры состоит из головной лопасти (простомиума), нескольких сегментов и маленькой анальной лопасти (пигидиума) с порошицей на конце. Головная лопасть представляет собой почти неизменившееся переднее предротовое полушарие трохофоры, а анальная лопасть— самый нижний участок трохофоры, лежащий позади уровня первичных мезодермальных клеток. Таким образом, оба отдела метатрохофоры по природе резко отличны от сегментов туловища. Существенная 264 особенность сегментов метатрохофоры заключается в том, что все они возникают одновременно. 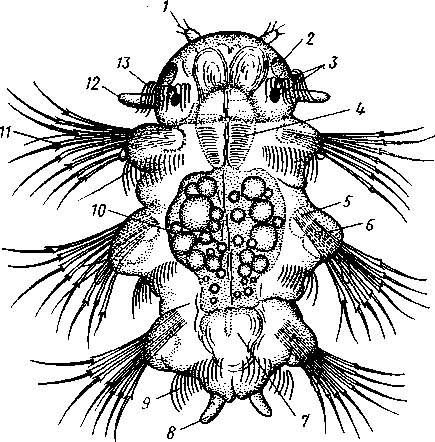 Рис. 226. Метатрохофора Nereis pelagica (no Рунштрому): / — щупальце. 2 — пигментная клетка. 3—глаза, 4 — глотка, 5 — щетинконосный мешочек параподии, 6 — па-раподия, 7 — задняя кишка, 8 — анальный усик, 9 — ресничный поясок, 10 — средняя кишка, 11 —щетинки, 12 — пальп, 13 — прототрох   Рис. 227. Схема соотношений между мезодермой лар-вальных и постларвальных сегментов при развитии Polychaeta (по П. Иванову). А — превращение тро-хофоры в мстатрохофору, мезодермальные полоски не сегментированы; Б — Метатрохофора с тремя лар-вальными сегментами, мезодермальные полоски сегментированы; В — появление на заднем конце тела эктодермальной зоны роста; Г — образование постларвальных сегментов из зоны роста (зона роста я целомические мешки постларвальных сегментов зачернены) Метатрохофора некоторое время плавает или ведет дойный образ жизни, не меняясь существенно, а затем испытывает дальнейший метаморфоз. На переднем крае анальной лопасти образуется зона роста, клетки которой непрерывно размножаются. Область, лежащая впереди нее, состоит из быстро растущих, еще не дифференцированных тканей. В этой зоне формируются новые сегменты и последовательно один за другим отделяются по направлению кпереди (рис. 227). Процесс продолжается до тех пор, пока не образуется столько сегментов, сколько их у взрослого червя. Иногда первичные мезодермальные клетки сохраняются в зоне роста и дают начало мезодермальным полоскам. Чаше, однако, эти клетки целиком расходуются на образование ие-ломической мезодермы метатрохофоры, и мезодермальные полоски образуются за счет размножения эктодермальных клеток зоны роста. От ме-зодермальных полосок последовательно отделяются парные зачатки цело-мических мешков (рис. 227). Каждый вновь образующийся сегмент получает свою пару таких зачатков, из которых развиваются его целомические мешки, разрастающиеся по бокам кишечника кверху и книзу, постепенно вытесняя собой первичную полость тела. Наконец, правый и левый мешки в каждом сегменте встречаются над и под кишкой по спинной и брюшной линиям, а следующие друг за другом пары мешков тоже соприкасаются своими передними и задними стенками 265 (рис. 228). Наружным кожно-мускульным листком мешки подстилают эктодерму, а внутренним граничат с кишечником. В результате описанного процесса первичная полость тела личинки замещается целомом. За счет соприкасающихся стенок целомических мешков над и под кишкой образуется спинная и брюшная брыжейки, а на месте соприкосно- 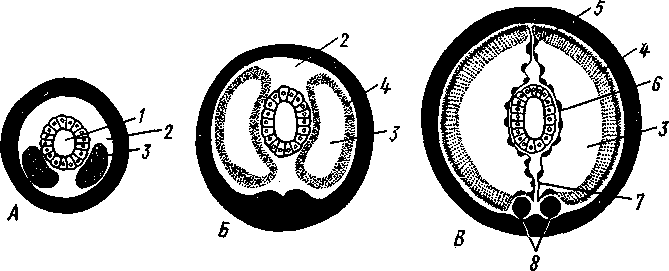 Рис. 228. Развитие целома у кольчатых червей. А—В — поперечные разрезы трех последовательных стадий развития сегмента (из Матвеева): / — кишка. 2 — первичная полость тела. 3 — целом, 4 — наружная стенка целомиче- ского мешка, 5 — спинной мезентерий, 6 — внутренняя стенка целомического мешка, 7— брюшной мезентерий, 8 — брюшные нервные стволы вения следующих друг за другом пар мешков — поперечные септы между полостями отдельных сегментов — диссепименты. Продольные кровеносные сосуды — спинной и брюшной — образуются за счет оставшихся участков первичной полости тела внутри брыжеек (между двумя сло-•ями перитонеального эпителия). Производные мезодермы таковы: из кожно-мускульного листка целомических мешков происходит мускулатура тела, из кишечно-мускульно-го — мускулатура кишечника. Кроме того, за счет стенок мешков формируются перитонеальный эпителий и половые воронки (целомодукты). На каждом сегменте развиваются параподии, а внутри него формируются соответствующие участки брюшных нервных стволов, брюшные ганглии, органы выделения и т. п. Важным для понимания организации кольчатых червей, так же как и других типов, берущих от них начало, является двойственность сегментов, входящих в состав тела этих животных. Мы видим, что в процессе метаморфоза сперва образуются сегменты метатрохофоры, затем формируются все остальные (см. рис. 227). Это явление было открыто крупнейшим советским эмбиологом П. П. Ивановым, разработавшим стройную теорию двойственного происхождения метамерии, имеющую большое значение для понимания эволюции всех сегментированных животных. Немногочисленные сегменты метатрохофоры названы П. П. Ивановым ларвальными (т. е. личиночными) в отличие от образующихся позднее постларвальных сегментов. Ларвальные сегменты характеризуются рядом существенных особенностей: образуются одновременно, причем сегментация начинается с наружных органов, отсутствуют половые железы и половые воронки, а иногда отсутствует и метамерность целома. Напротив, при образовании постларвальных сегментов из зоны роста прежде всего сегментируется мезодерма, образуются кровеносная система, половые железы и половые воронки. 266 Таким образом, тело взрослого полимерного червя состоит из следующих различных по происхождению отделов: 1) головной лопасти, или простомиума, представляющего видоизмененное переднее полушарие трохофоры; 2) нескольких ларвальных сегментов; 3) многочисленных постларвальных сегментов и 4) анальной лопасти, или пигидиума, происходящей из самого заднего участка трохофорной личинки. Олигомерные кольчецы (Dinophilus, Myzostornum и др.) в составе туловища имеют только ларвальные сегменты. Поэтому они во многих отношениях напоминают личинку — метатрохофору. Итак, важнейшие черты развития полихет заключаются в спиральном, детерминативном типе дробления, в образовании личинки трохофоры и в закладке мезодермы путем отделения двух мезодермальных полосок от двух мезобластов — первичных мезодермальных клеток; эти клетки называют телобластами, соответственно и данный способ закладки мезодермы именуется телобластическим. Экология. К классу Polychaeta принадлежит около 5300 видов, главным образом морских. Только немногие представители встречаются в пресноводных бассейнах, например в Байкале (Manayunkia). Большинство многощетинковых ведет донный образ жизни, встречаясь главным образом в прибрежной полосе. Многие из них спускаются, однако, глубже 1000 м, а некоторые были найдены даже на глубине 8000 м. Сравнительно немногие виды ведут свободноплавающий образ жизни в планктоне (сем. Alciopidae и др.) и, подобно многим другим планктонным животным, отличаются стекловидной прозрачностью тела. Донные многощетинковые большей частью ползают по дну, среди водорослей, но многие из них ведут роющий образ жизни, проделывая в песке или в иле длинные норы; таков крупный морской червь пескожил — Arenicola (см. рис. 211, Г) и др. Особую биологическую группу составляют сидячие полихеты, выделяющие вокруг себя защитные трубки, из которых высовывается лишь передний конец червя. Размеры представителей класса Polychaeta колеблются в пределах от нескольких миллиметров до 3 м (Eunice gigantea). Палеонтология. Ископаемые остатки полихет немногочисленны. Чаще всего сохраняются трубки сидячих форм, известные еще с докембрия. От бродячих полихет остаются отпечатки щетинок и челюстей, как, например, у Eunicites из литографского сланца Баварии. Практическое значение. Некоторые тропические формы вроде упомянутого выше палоло—Eunice viridis (см. рис. 222) употребляются человеком в пищу. Отдельные виды используются в качестве наживки при ловле рыбы, например пескожил (Arenicola marina', см. рис. 211,Г). Донные формы многощетинковых червей в больших количествах поедаются промысловыми рыбами, камчатским крабом и другими животными. В настоящее время основу пищевого рациона осетровых рыб Каспийского моря составляют Nereis diversicolor— донный многощ-етинковый червь, который отсутствовал в Каспии, но в 1939 и 1940 гг. был переселен туда из Азовского моря. Исследования, связанные с этим успешным переселением ценного кормового объекта, были осуществлены под руководством крупнейшего советского гидробиолога академика Л. А. Зенкевича. Классификация. Класс Polychaeta делится на два подкласса: бродячие (Errantia) и сидячие (Sedentaria). ПОДКЛАСС I. БРОДЯЧИЕ (ERRANTIA) Головная лопасть хорошо развита, сегменты более или менее гомо-номны. Параподии хорошо развиты на всем протяжении тела, часто снабжены жабрами. Нефридии имеют метамерное расположение. В большинстве — свободноподвижные хищники. Представители: Aphrodite, или «морская мышь», густо покрытая длинными щетинками, Lepidonolus (см. рис. 211, Б), у которого тело покрыто двумя рядами пластинок — элитр, представляющих видоизмененные усики параподий, Alciope v ana-dis — планктонные прозрачные полихеты с сильно развитыми глазами, Nereis (см. рис. 211, А) —обыкновенная форма, обладающая эпитокной половозрелой стадией. С подклассом Errantia, по-видимому, связана небольшая группа Му-zostomida, которую можно считать одним из отрядов бродячих полихет. Это сильно укороченные и уплощенные животные всего с пятью парами слабо развитых параподий. Myzostomida паразитируют на или внутри иглокожих, главным образом на морских лилиях. К подклассу Errantia относятся и роющиеся в грунте Polygordius, Protodrilus и другие близкие к ним формы, а также олигомерный Dinop-hilus (см. рис. 212). Иногда на основании очень большой примитивности организации их выделяют в самостоятельный класс первичных кольчецов (Archiannelida). ПОДКЛАСС П. СИДЯЧИЕ (SEDENTARIA) Головная лопасть слабо развита или редуцирована. Тело часто поделено на несколько гетерономных отделов, параподий развиты слабо, жабры ограничиваются лишь известным участком тела, чаще всего головным. Нефридии развиты далеко не во всех сегментах. Живут обыкновенно внутри трубок, постоянных или временных. Представители: Chae-topterus — тело гетерономно расчлененное, обладает способностью ярко светиться в темноте, Arenicola (см. рис. 211, Г) —пескожил, достигающий в длину свыше 30 см, делает норки в песке, Serpula и Spirorbis (см. рис. 211, Д, Ж) — обладают сильно разветвленными пальпами и живут в известковой трубке, вход в которую животное может закрывать особой крышечкой, представляющей видоизмененное щупальце. ПОДТИП II. ЛОЛСНОВЫЕ (CLITELLATA) Для представителей подтипа Clitellata характерен гермафродитизм. Гонады утрачивают метамерное расположение. Имеется поясковая зона (clitellum). Развитие прямое. К подтипу относятся два класса: Малоще-тинковые (Oligochaeta) и Пиявки (Hirudinea). |