Хрестоматия. Петухов. Том 3. Книга 2. Учебник по общей психологии, предназначено для проведения семинарских занятий по данному курсу и самостоятельного чтения
 Скачать 20.88 Mb. Скачать 20.88 Mb.
|

|
3. Восприятие движения. Системы восприятия реального движения: изображение/сетчатка и глаз/ голова. Теории восприятия стабильности видимого мира. Иллюзии восприятия движения: автокинетическое, стробоскопическое и индуцированное движение, эффект водопада. Восприятие времени Р. Грегори ЗРИТЕЛЬНОЕ ВОСПРИЯТИЕ ДВИЖЕНИЯ1 Восприятие движения имеет жизненно важное значение. Для животных, стоящих на эволюционной лестнице ниже человека, движущиеся объекты являются, вероятно, сигналами либо опасности, либо потенциальной пищи и требуют быстрого соответствующего действия, в то время как неподвижные объекты могут быть игнорированы. Фактически, вероятно, только глаза высших животных могут давать мозгу информацию о неподвижных объектах. Некоторые особенности эволюционного развития зрительной системы, начиная от глаза, способного воспринимать лишь движения, и кончая глазом, воспринимающим формы, сохранились в строении сетчатки человеческого глаза. Края сетчатки чувствительны только к движению. Это можно видеть, совершая колебательные движения каким-либо предметом в области периферии зрительного поля так, чтобы стимулировались только края сетчатки. Вы увидите, что при этом воспринимается только движение и его направление, но невозможно определить, какой предмет движется. Это очень близко к тому, что наблюдается при примитивном восприятии. Самые периферические отделы сетчатки еще более элементарны; когда они стимулируются движениями, мы еще ничего не воспринимаем, однако эта стимуляция вызывает рефлекс поворота глаз, благодаря которому изображение объекта перемещается в центральное поле зрения, с тем чтобы наиболее высоко организованная фо-веальная область сетчатки с ее объединенными в нервную сеть элементами приняла участие в опознании объекта. Таким образом, периферия сетчатки представляет собой аппарат для раннего обнаружения объекта, он вызывает поворот глаз для того, чтобы цель попала на объекторазличительную часть системы, оценивающую объект как полезный, вредный или нейтральный. 1 Грегори Р. Глаз и мозг. М.: Прогресс, 1970. С. 101-130. 198 Тема 17. Экспериментальные исследования восприятия Р 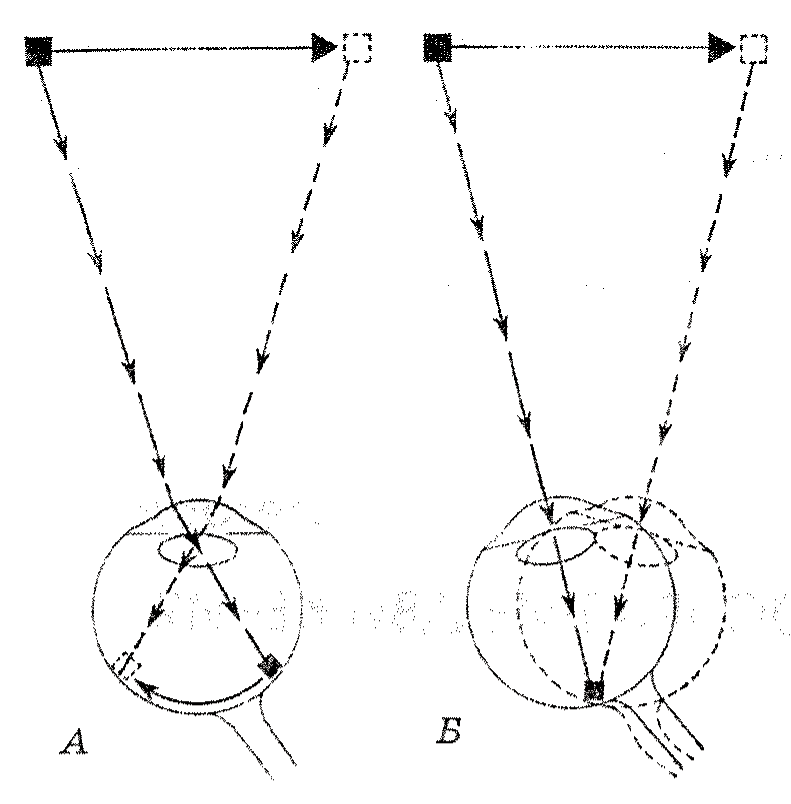 ис. 1. А - система восприятия движения изображение/сетчатка: изображение движущегося объекта пробегает по сетчатке в то время, когда сами глаза остаются неподвижными; таким образом, информация о движении возникает путем последовательной стимуляции рецепторов в соответствии с траекторией движения объекта; Б - система восприятия движения глаз/голова: когда глаз следует за движущимся объектом, изображение остается стационарным на сетчатке, но мы продолжаем видеть движение. Эти две системы иногда могут давать противоречивые показания, что приводит к любопытным иллюзиям ис. 1. А - система восприятия движения изображение/сетчатка: изображение движущегося объекта пробегает по сетчатке в то время, когда сами глаза остаются неподвижными; таким образом, информация о движении возникает путем последовательной стимуляции рецепторов в соответствии с траекторией движения объекта; Б - система восприятия движения глаз/голова: когда глаз следует за движущимся объектом, изображение остается стационарным на сетчатке, но мы продолжаем видеть движение. Эти две системы иногда могут давать противоречивые показания, что приводит к любопытным иллюзиямТакие глаза, как наши собственные, подвижные относительно головы, могут давать информацию о движении двумя различными способами. Когда глаз остается неподвижным, образ движущегося объекта перемещается по рецепторам сетчатки и вызывает в них быстро сменяющиеся сигналы; но когда сам глаз следует за движущимся объектом, его изображение остается более или менее неподвижным относительно сетчатки, так что оно не может быть сигналом движения, однако мы все же видим движение объекта. Если объект воспринимается на неподвижном фоне, быстро сменяющиеся сигналы могут возникать теперь от фона, который передвигается по сетчатке во время слежения глаз за движущимся объектом; однако, мы продолжаем видеть движение даже при отсутствии фона. Это можно показать на простом опыте. Попросите кого-нибудь медленно помахивать зажженной сигаретой в темной комнате и последите за ней глазами. Движение сигареты видно, хотя в данном случае нет сигналов фона, двигающихся по сетчатке. Очевидно, повороты глаз относительно головы могут дать восприятие движения и довольно точную оценку скорости движения и при отсутствии сигналов, передвигающихся по сетчатке. Следовательно, существуют две системы восприятия движения; мы назовем одну из них (А) система изображение/сетчатка; другую (В) система глаз/голова (рис. 1). (Эти названия заимствованы из артиллерийского дела, где возникают сходные ситуации, когда орудие нацеливается на объект с движущейся палубы корабля. Орудийная башня может быть неподвижна или следовать за целью, но движение цели в каждом случае может быть обнаружено.) Рассмотрим теперь систему изображение/сетчатка, а затем обратимся к тому, как эти две системы работают совместно. Грегори Р. Зрительное восприятие движения 199 Система восприятия движения изображение/сетчатка С помощью регистрации электрической активности сетчатки глаз животных было обнаружено, что существуют различного рода рецепторы, подавляющее большинство которых сигнализирует только об изменении освещенности, и только немногие отвечают длительным возбуждением на постоянный свет. Некоторые рецепторы возбуждаются при включения света, другие — при его выключении, третьи — как при включении, так и при выключении. Эти различного рода рецепторы сетчатки названы соответственно рецепторами «включения», рецепторами «выключения» и рецепторами «включения — выключения». По-видимому, эти рецепторы, чувствительные только к изменениям освещения, и ответственны за сигнализацию движения; таким образом, все глаза являются прежде всего детекторами движения. Эти рецепторы, сигнализирующие только об изменении освещенности, будут отвечать на движущиеся края изображения, но не будут реагировать на неподвижные изображения до тех пор, пока сами глаза не начнут двигаться. С помощью тонких проволочных электродов, помещенных на сетчатку изолированного глаза лягушки, было обнаружено, что анализ ре-цепторной активности происходит в сетчатке задолго до того, как сигналы достигнут мозга. В статье с интригующим названием «Что глаз лягушки сообщает мозгу лягушки», написанной Летвином, Матураной, МакКеллоком и Питсом из лаборатории электроники Массачусетского технологического института, сетчатка описывается как «детектор насекомых»; авторы обнаружили три класса волокон, посылающих в мозг различного рода информацию. «Детектор насекомых» вызывает рефлекс движения языком, когда на сетчатку падает маленькая тень, отбрасываемая, например, мухой; таким образом, сетчатка в данном случае функционирует как мозг. Кроме этой системы, которая отвечает, по существу, на кривые линии, они обнаружили:
Глаз лягушки сигнализирует только об изменении освещенности и движении изогнутых краев объектов; все остальное игнорируется и никогда не доходит до мозга. Зрительный мир лягушки, таким образом, ограничен лишь движением некоторых видов объектов. Физиологи Хьюбел и Визел провели важное исследование, регистрируя электрическую активность зрительной области мозга кошки. Они обнаружили, что в ней существуют отдельные клетки, которые отвечают 200 Тема 17. Экспериментальные исследования восприятия только на движение изображения по сетчатке, причем на движение, осуществляемое только в одном определенном направлении. <...> Тот факт, что движение перекодируется в нервную активность сетчатки или в активность зрительных проекционных областей мозга, находящихся непосредственно за сетчаткой, представляет собой физиологическое открытие, важное со многих точек зрения, и прежде всего потому, что оно показывает, что скорость движения может восприниматься независимо от оценки времени. Однако часто считают, что нервная система, ответственная за восприятие скорости движения, должна представлять собою своего рода «внутренние часы». Скорость в физике определяется как время, необходимое для того, чтобы объект переместился на определенное расстояние (v=d/t). Следовательно, предполагается, что для оценки скорости движения всегда необходима оценка времени. Но ведь спидометр автомобиля не имеет в своем устройстве часов. Часы нужны для калибровки этого прибора после его изготовления, но однажды откалиброванный, он будет измерять скорость движения без часов; то же самое справедливо, вероятно, и по отношению к глазу. Изображение, пробегающее по сетчатке, последовательно возбуждает рецепторы, и чем быстрее это изображение движется, тем — до известных пределов — более интенсивные сигналы скорости оно вызывает. Аналогия с другими измерителями скорости (спидометром и т.п.) показывает, что скорость может быть оценена безотносительно к «часам», но эта аналогия еще не говорит нам точно, как работает при этом нервная система. Когда-нибудь будет возможно изобразить полную круговую схему сетчатки и создать ее действующую электронную модель; однако пока мы не можем сделать это с полной уверенностью в отношении человеческого глаза. Такая модель была предложена для фасеточного глаза жука. Эта модель была изготовлена, и теперь она иногда используется в воздушном флоте, чтобы определять отклонение самолета от курса под влиянием ветра. Глаз как детектор движения сформировался в процессе биологической эволюции несколько сот миллионов лет тому назад, принцип его действия раскрыт с помощью электроники, а затем был построен его электронный эквивалент, который теперь используется при полетах человека. Система восприятия движения глаз/голова Нервные аппараты, обеспечивающие восприятие движения посредством перемещения изображения по сетчатке, существенно отличаются от другого способа сигнализации движений с помощью поворота глаза. Каждый глаз имеет шесть внешних мышц, управляющих его движениями; любое движение глаз сигнализируется в мозг и используется в качестве индикатора движения внешних объектов. То, что это действительно Грегори Р. Зрительное восприятие движения 201 так, показывает опыт с сигаретой, который мы уже описывали; в этом случае нет никакого систематического движения изображения по сетчатке, и тем не менее движение сигареты, прослеживаемое глазами, видно (рис. 1, Б). Самым вероятным типом сигналов, возникающих при этом, были бы обратные сигналы от мускулатуры глаза, так что, когда происходит растяжение мышц глаза, в мозг посылаются обратные сигналы, указывающие на движение глаз, а также объектов, прослеживаемых взором. Таково было бы инженерное решение этой проблемы, но так ли решает ее природа? Мы можем получить ответ, если займемся, казалось бы, совсем иным вопросом. Почему мир остается стабильным, когда наши глаза двигаются? Сетчаточные изображения перемещаются по рецепторам сетчатки всякий раз, когда наши глаза двигаются, — и все же мы не воспринимаем движения, мир не вращается, как бы наши глаза ни двигались. Почему это так? Как мы знаем, существуют две нервные системы сигнализации движений: система изображение/сетчатка и система глаз/голова. Очевидно, во время нормальных движений глаз эти системы тормозят друг друга, в результате чего и возникает стабильность зрительного мира. Идея взаимного торможения этих систем как средства стабилизации зрительного восприятия рассматривалась Чарлзом Шеррингтоном - физиологом, внесшим значительный вклад в анализ спинальных рефлексов, а также Гельм-гольцем; однако они объясняли это явление с различных позиций и особенно расходились в оценке деятельности той системы, которую мы называем системой восприятия скорости движения глаз/голова. Теория Шеррингтона известна под названием афферентной теории, а Гельмголь-ца - под названием эфферентной теории (рис. 2). Шеррингтон думал, что сигналы от глазных мышц составляют систему обратных афферентаций, поступающих в мозг, когда глаза двигаются, и что они тормозят сигналы движения, возникающие в сетчатке. Это представление известно в технике как обратная связь; однако для нервных сигналов, поступающих от глазных мышц, требуется довольно длительное время, чтобы дойти до мозга, и, если принять эту точку зрения, следовало бы ожидать появления неприятных ощущений неустойчивости всех видимых предметов каждый раз, когда мы двигаем глазами, до тех пор пока афферентные сигналы от глазных мышц не достигнут мозга и не затормозят сетчаточных сигналов движения. Гельмгольц высказал совершенно иное предположение. Он считал, что сетчаточные сигналы движения тормозятся не сигналами от глазных мышц, а центральными сигналами, исходящими от мозга и управляющими самими движениями глаз. 202 Тема 17. Экспериментальные исследования восприятия 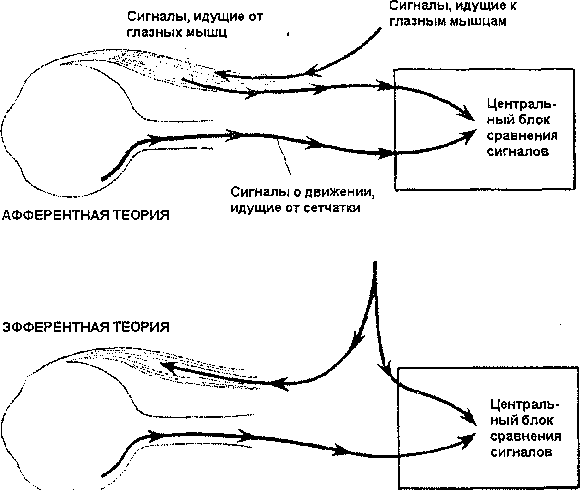 Рис. 2. Почему мир остается стабильным, когда наши глаза двигаются? Согласно афферентной теории, сигналы движения, поступающие от сетчатки (от системы изображение/сетчатка), тормозятся сигналами, идущими от глазных мышц (афферентными). Согласно эфферентной теории, сетчаточные сигналы движения тормозятся сигналами команды, управляющими самими движениями глаз, сигналами (эфферентными), которые, в свою очередь, регулируются внутренней замкнутой системой мозга. Факты свидетельствуют в пользу эфферентной теории Решение этого вопроса может быть получено с помощью очень простых экспериментов, которые читатель может проделать на себе самом. Попробуйте осторожно двигать глаз пальцем, закрыв другой глаз рукой. Когда глаз смещается пассивно, мир будет казаться вращающимся в направлении, противоположном движению глаза. Очевидно, стабильность видимого мира поддерживается не пассивными, а нормальными произвольными движениями глаз. Так как мир движется в направлении, обратном направлению пассивного движения глаза, очевидно, что система восприятия движения изображение/сетчатка продолжает работать; здесь выключена только система глаз/голова. Можно было бы спросить, почему система глаз/голова связана только с произвольными, но не с пассивными движениями глаз? Шеррингтон полагал, что эта система работает с помощью сигналов, идущих от рецепторов растяжения, находящихся в глазных мышцах. Такие рецепторы растяжения мышц хорошо известны, они посылают обратные сигналы от мускулатуры при движении ко- Грегори Р. Зрительное восприятие движения 203 нечностей. Однако создается впечатление, что система восприятия движения глаз/голова работает иным образом, так как рецепторы растяжения продолжают посылать сигналы и при пассивном состоянии глазных мышц. Мы можем прекратить все сетчаточные сигналы движения и посмотреть, что произойдет при пассивном перемещении глаза. Это можно легко сделать с помощью засвета ярким светом (или фотографической вспышкой), чтобы получить последовательный образ. Это вызовет утомление одного определенного места сетчатки, соответствующего фотографической вспышке, и этот образ будет передвигаться точно вместе с глазом, так что, хотя глаз и будет двигаться, сигналы от перемещения изображений по сетчатке не смогут возникнуть. Если мы будем наблюдать за последовательным образом в темноте (чтобы избежать фона), мы обнаружим, что, когда глаза пассивно приводятся в движение пальцем, последовательный образ не перемещается. Это очень убедительный довод против афферентной теории, так как активность рецепторов растяжения должна была бы вызвать перемещение последовательного образа вместе с глазом, если бы эта активность в обычных условиях тормозила сетчаточные сигналы движения. Теперь, если глаз будет двигаться произвольно, мы обнаружим, что последовательный образ перемещается вместе с глазом. Куда бы глаз ни переместился, последовательный образ будет следовать за ним. Гельмгольц при объяснении этого факта исходил из предположения, что здесь мы имеем дело не с афферентной активностью, идущей от глазных мышц, вовлеченных в движение, а с эфферентными сигналами команды, управляющими движением глаз. Эта эфферентная теория, как мы уже видели, утверждает, что сигналы команды регулируются внутренней замкнутой системой мозга и подавляются сетчаточными сигналами движения. Когда этих сет-чаточных сигналов нет, как в случае с последовательным образом, видимым в темноте, мир вращается вместе с глазом, потому что сигналы команды не тормозятся сетчаткой. Пассивные движения глаза не вызывают движения последовательного образа, так как в этом случае нет системы, которая давала бы сигналы движения. В клинических случаях, при каких-либо нарушениях глазных мышц или их нервного аппарата, у пациентов появляется ощущение вращения окружающих предметов, когда они пытаются двигать глазами. Их мир движется в том же направлении, в котором они намеревались двигать глазами. Это происходит также и тогда, когда мышцы глаза парализуются с помощью кураре — южноамериканского яда для стрел. Немецкий ученый Эрнст Мах фиксировал свои глаза мастикой так, что они не могли двигаться, и он получил те же результаты. Система глаз/голова, таким образом, приводится в действие не фактическими движениями глаз, а командой двигать глазами. Она работает даже в тех случаях, когда глаза не повинуются команде. Удивительно, 204 Тема 17. Экспериментальные исследования восприятия что сигналы команды могут вызывать восприятие движения: принято думать, что восприятие движения исходит от глаз, а не от находящихся в глубине мозга аппаратов, контролирующих движения глаз. Почему же возникла такая странная система? Это тем более удивительно, что в глазных мышцах действительно были обнаружены рецепторы растяжения. Афферентная система, или система обратных связей, по-видимому, действовала бы слишком медленно: пока сигнал обратной связи достиг бы мозга, чтобы затормозить сетчаточные сигналы движения, было бы слишком поздно. Тормозящий сигнал мог бы начаться в тот же самый момент, что и команда к движению глаз, и тогда он мог бы затормозить сетчаточный сигнал без опоздания. Действительно, для того, чтобы сигнал сетчатки достиг мозга, требуется немного времени («время сетчаточной реакции»), но тогда сигнал команды пришел бы в мозг для затормаживания сетчаточного сигнала слишком рано, однако этот сигнал команды задерживается, чтобы совпасть по времени с сигналом сетчатки. В этом мы можем убедиться при тщательном исследовании движения последовательного образа при произвольных движениях глаз. Всякий раз, когда глаз двигается, требуется некоторое время, чтобы возникло движение последовательного образа, и, очевидно, эта отсрочка и приводит к тому, что управляющий командный сигнал достигает мозга не раньше, чем сигнал от сетчатки. Можно ли представить себе более совершенную систему? |