|
|
этик. Учебное пособие для студентов лечебного, педиатрического, стоматологического и фармацевтического факультетов Тверь, 2006 г Лекции Гормоны и гормоноподобные вещества
Липофильные гормоны: Это прежде всего стероидные гормоны коры надпочечников – глюко- и минералокортикоиды, а также половые гормоны: женские – эстрогены и прогестины, мужские – андрогены.
К группе липофильных относят также гормоны щитовидной железы (Т3, Т4).
Стероидные гормоны
Е1…Х
Е1…
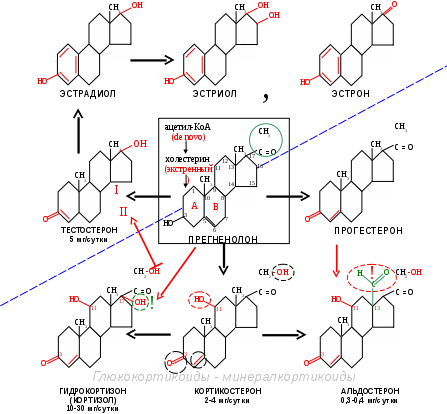
Биосинтез и распад стероидных гормонов. Все стероидные гормоны имеют в основе структуру циклопентанпергидрофенантрена, поэтому общим предшественником всех стероидных гормонов является холестерин (С27), который поступает в гормонсинтезирующие клетки в
составе ЛПНП или синтезируется в них из ацетил-КоА. Катализируют образование стероидных гормонов из холестерина высокоспецифические ферменты. Биосинтез каждого стероидного гормона состоит из множества последовательных ферментативных реакций. Он начинается с удаления от холестерина части боковой цепи, которое приводит к образованию общего предшественника всех стероидных гормонов – прегненолона. Затем осуществляются у разных гормонов реакции гидроксилирования, изомеризации, гидратирования (+Н) или дегидратирования
(- Н).
Отличаются все стероидные гормоны по числу оставшихся углеродных атомов, типу заместителей и положению двойных связей:
холестерин и кальцитриол является С27-стероидами;
прогестерон, кортизол и альдостерон – С21-стероидами;
тестостерон является С19-стероидом;
эстрогены являются С18-стероидами.
Выполнив функции (речь о которых ниже), стероидные гормоны инактивируются. Механизм их инактивации – ферментативный процесс, который происходит в печени. Молекулы стероидных гормонов коры надпочечников и тестостерон подвергаются расщеплению до 17- кетостероидов (С17), эстрогены восстанавливаются по месту двойной связи и подвергаются гидроксилированию, а затем они конъюгируются с серной или глюкуроной кислотой и удаляются из организма с мочой или частично с желчью. В связи с этим, определение в моче конечных продуктов их инактивации используется для характеристики функциональной активности эндокринных желез.
Гормоны коры надпочечников
Кора надпочечников взрослого человека состоит их трех слоев. Субкапсулярная область, называется клубочковой зоной – она продуцирует минералокортикоиды. В пучковой зоне и в следующей за ней сетчатой зоне вырабатываются глюкокортикоиды и андрогены. Андрогены надпочечников служат основными предшественниками эстрогенов у женщин в постменопаузе.
Стимулирует их образование АКТГ. Прикрепляясь к рецепторам клеток коры надпочечников, АКТГ усиливают образование ц-АМФ. Это приводит к активации эстераз и образованию из эфиров холестерина свободного холестерина, который транспортируется в митохондрии, где фермент цитохром Р450 отщепляет боковую цепь и превращает его в прегненолон. Биосинтез гормонов может начинаться также с ацетил-КоА, но и в этом случае вначале образуется холестерин. Дальнейшие реакции катализируются дегидрогеназами, лиазами, изомеразами. В результате из прегненолона образуются: дезоксикортикостерон и кортикостерон (2-4 мг/сутки), а через ряд других химических реакций альдостерон (0,3-0,4 мг/сутки).
Для синтеза кортизола (гидрокортизона), самого мощного из природных глюкокортикоидных гормонов (10-30 мг/сутки) человека, необходимо еще одно гидроксилирование (см. схему).
Стероидные гормоны практически не накапливаются в клетках надпочечников, а высвобождаются в плазму крови по мере их образования.
Транспорт в крови. Кортизол, кортикостерон и дезоксикортикостерон не растворимы в воде и потому в плазме крови находятся в форме преимущественно связанной с белками, и лишь небольшая часть - в свободном виде. Связывающий гормон белок плазмы α - глобулин называют транскортином (кортикостероид - связывающий белок). Образуется этот белок в печени. От прочности связи гормона с белком зависит длительность периода полужизни этих гормонов. Так, у кортизола, связанного с транскортином прочно, t1/2 = 1,5-2,0 часа, а связанный слабо - 1 час. Связанный с белком гормон неактивен. Несвязанный (свободный) кортизол составляет 8% от общего количества гормона в плазме и представляет активную фракцию.
Альдостерон не имеет специфического транспортного белка в плазме и связывается с альбумином.
Скорость секреции кортизола (глюкокортикоидов) зависит от концентрации АКТГ (гормон гипофиза), выделение которого в свою очередь контролируется выделением кортиколиберина (гормон гипоталамуса), т.е. эти гормоны связаны между собой классическими петлями прямой и отрицательными обратными связями.
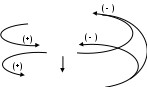
гипоталамус
(кортиколиберин)
гипофиз
(АКТГ)
кора надпочечников
(глюкокортикоиды)
( + ) Кортиколиберин усиливает синтез и выделение АКТГ гипофизом.
( + ) АКТГ повышает скорость отщепления боковой цепи от холестерола и усиливает образование кортизола в коре надпочечников.
( - ) По мере повышения в крови уровня глюкокортикоидов (кортизола) происходит снижение секреции кортиколиберина, что ведет к снижению выработки АКТГ гипофизом, а затем и кортизола надпочечниками.
Помимо того, высвобождение АКТГ и секреция кортизола регулируются нервными импульсами, поступающими из разных отделов нервной системы. Этими механизмами формируется так называемый эндогенный ритм синтеза гормонов коры надпочечников. Уровень кортизола начинает возрастать во время сна, достигая пика вскоре после пробуждения, затем уровень гормонов постепенно падает до минимальных величин к концу дня. Эти колебания в образовании и содержании глюкокортикоидов зависят от светового периода, циклов (питания – голодания), (сна – бодрствования). Исчезновение суточной периодичности секреции стероидов обычно связано с патологией гипофизарно-адреналовой системы, некоторыми видами депрессивных состояний. На секрецию кортизола оказывают также сильное влияние физический и эмоциональный стрессы, состояния тревоги, страха, волнения и боль. Эти реакции могут нарушать действие системы отрицательной обратной связи и суточного ритма производства глюкокортикоидных гормонов.
Продукция альдостерона (минералокортикоида) клетками клубочковой зоны регулируется сложнее и совершенно иначе: основными регуляторами в этом случае служат система ренин-антиотензин-калий, а также участвуют в этом процессе Nа, АКТГ и нейрональные механизмы.
Система ренин-ангиотензин.
Эта система (совместно с альдостероном) участвует в регуляции кровяного давления, электролитного и водного баланса.
I. В печени синтезируется особый белок α 2 – глобулин – незрелый белок, который называют ангиотензиногеном. Он служит субстратом для фермента - ренина, продуцируемого юкстагломерулярными клетками почечных афферентных артериол. Локализация клеток ЮГА делает их особенно чувствительными к изменениям кровяного давления. Эти клетки чувствительны также к изменению концентрации Nа, К в жидкости, протекающей через почечные канальцы. В результате любая комбинация факторов вышеперечисленных изменений (снижения объема жидкости, снижение АД, снижение Na Cl и др.) стимулирует высвобождение ренина. На освобождение ренина оказывают также влияние ЦНС и изменения положения тела. Соответствующие сигналы поступают по симпатическим нервам к юкстагломерулярным клеткам.
II. Освобождающийся в кровь из почек ренин действует на ангиотензиноген (
400 аминокислотных остатков) и превращает его в декапептид –
ангиотензин -I.
III. Далее на ангиотензин- I действует ангиотензин - превращающий фермент, синтезируемый в легких и эндотелиальных клетках. Фермент отщепляет 2 аминокислотных остатка и превращает ангиотензин - I в ангиотензин-II и III
(8-7-пептиды). Ангиотензин - II: - увеличивает кровяное давление, вызывая
1) сужение артериол, являясь самым сильнодействующим из известных вазоактивных агентов;
2) оказывает сильное стимулирующее действие на выработку альдостерона.
3) помимо того ангиотензин-превращающий фермент расщепляет также брадикинин - мощное сосудорасширяющее средство. Следовательно, ангиотензин - расщепляющий фермент повышает кровяное давление двумя различными способами, действуя на:
ангиотензин-I, превращая его в ангиотензин II
брадикинин, разрушая последний
1 ) почки ренин ) почки ренин
2) печень ангиотензиноген
3) легкие ангиотензин - ангиотензин-I
превращающий
фермент ангиотензин-II
 Na+ Na+
  K+ K+
2
брадикинин 1 усиление биосинтеза
(усиление распада) альдостерона
усиление реабсорбции
4) сосуды сужение Na+ в почках
кровеносных сосудов
усиление реабсорбции
Н2О в почках
увеличение объема крови
2 1
( ↑АД ) повышение кровяного давления
Влияние кортикостероидных гормонов на обмен веществ и физиологические функции
Кортикостероиды оказывают многообразное влияние на обмен веществ и физиологические функции:
Оказывают выраженное влияние на воспалительные и аллергические реакции;
Контролируют систему кроветворения и иммунитет;
Повышают устойчивость организма к повреждающим факторам и обеспечивают сохранение гомеостаза;
Регулируют обмены углеводов, липидов, белков и нуклеиновых кислот;
Регулируют обмен электролитов и воды, включая циркуляторный гомеостаз и нервно-мышечную возбудимость;
Оказывают выраженное секреторное действие (усиливая секрецию HCL и энзимов переваривания белков в желудочно-кишечном тракте).
Избыточное или недостаточное содержание в крови глюко- и минералокортикоидов вызывает ряд серьезных осложнений в обмене веществ и физиологических функциях. Утрата кортикоидной функции надпочечников (в отсутствии заместительной терапии) ведет к летальному исходу. Лечение глюкокортикоидами и минералокортикоидами не дает должного эффекта.
Важно отметить, и это должен помнить каждый врач, что при введении любого из трех главных гормонов коры надпочечников, (глюко- минерал- кортикоидов), а именно кортизола (гидрокортизона), кортикостерона или альдостерона наблюдаются в общем сходные ответные реакции, перечисленные в пунктах (1-6). Однако в эффекте действия кортизола, с одной стороны, и альдостерона, с другой на углеводный и минеральный обмен наблюдаются крайние позиции, а кортикостерон занимает как бы промежуточное между ними положение:
кортизол (гидрокортизон) оказывает мощное влияние на обмен углеводов и характеризуется относительно слабым влиянием на обмен воды и минеральных веществ.
альдостерон, наоборот, оказывает сильное влияние на обмен воды и электролитов, по другим же показателям его эффективность составляет около 1/3 от активности кортизола.
В связи с этим кортизол и кортикостерон стали называть глюкокортикоидными гормонами (ГКГ), а альдостерон – минералокортикоидом (МКГ). Однако, врач должен помнить о широком диапозоне действия на обмен веществ и физиологические функции каждого из названных гормонов, чтобы не вызвать у пациента при гормонотерапии нежелательных осложнений, речь о которых будет ниже.
Широкое применение в клинике глюкокортикоидных гормонов и их искусственных аналогов объясняется, прежде всего, их выраженным влиянием на воспалительные и аллергические реакции, иммунитет, а также повышением устойчивости организма к различным повреждающим факторам (гомеостаз).
Противовоспалительный и антиаллергический эффект глюкортикоидных гормонов: объясняется тем, что они:
подавляют освобождение арахидоновой кислоты из состава фосфолипидов и ограничивают биосинтез из них простагландинов, стимулирующих воспалительный процесс. Кстати, широко известный аспирин подавляет активность простагландин-синтетазы и потому также оказывает жаропонижающий и противовоспалительный эффект.
тормозят накопление лейкоцитов в участке воспаления и формирование некроза тканей
ингибируют пролиферацию фибробластов и образование ими в очаге воспаления избытка коллагена и фибронектина, которые затрудняют кровообращение и очищение поврежденной ткани от накопления продуктов некроза, предотвращая тем самым формирование рубцовой ткани
Влияние ГКГ на систему кроветворения и иммунитет
При введении избытка ГКГ развивается лимфопения. Она обусловлена инволюцией лимфоидной ткани. В результате понижаются: пролиферация В-лимфоцитов в ответ на действие антигенов и выработка ими антител; а также супрессорные и хелперные функции Т-лимфоцитов. Эти реакции организма на введение ГКГ лежат в основе их использования при лечении аутоиммунных заболеваний и подавления реакции отторжения тканей при их пересадке (почки, печень, миокард, легкие). ГКГ оказывают также эритропоэтическое действие на костный мозг и повышают в крови содержание эритроцитов.
Влияние ГКГ на устойчивость организма к повреждающим факторам.
При недостаточности активности коры надпочечников у больных резко понижается возможность сохранения гомеостаза при действии на него различных повреждающих факторов, а именно понижается устойчивость к геморрагии (кровотечению), физической травме, инфекционным и сенсибилизирующим агентам, действию вредных химикалий. Кортизол, помимо того тормозит проявление повышенной реактивности организма, особенно при анафилактическом шоке. Если при травме или хирургической операции понижается секреция кортизола, то у пациента резко ослабевают шансы на выживание. В этих случаях уместна заместительная терапия глюкокортикоидами.
Влияние ГКГ на обмен углеводов, липидов, белков и нуклеиновых кислот
Столь разнообразное влияние ГКГ на обмен веществ объясняется тем, что эти гормоны изменяют (чаще повышают) в клетке содержание многих ферментов, действуя на ДНК, их регуляторные гены, стимулируя образование м-РНК и биосинтез белков-энзимов.
Влияние ГКГ на обмен углеводов. Они повышают образование глюкозы в печени посредством:
1) мобилизации аминокислот из периферических тканей путем усиления катаболизма белков и использования их в реакциях глюконеогенеза;
увеличения скорости глюконеогенеза путем повышения в клетке ключевых ферментов глюконеогенеза (ФЕПКК)
2) помимо того, ГКГ тормозят катаболизм глюкозы периферическими тканями. В итоге уровень глюкозы в крови повышается. Это может стать причиной гипергликемии, перенапряжения инсулярного аппарата при длительной терапии глюкокортикоидными гормонами и формирования стероидного диабета.
Влияние ГКГ на обмен липидов
Избыточное количество ГКГ стимулирует липогенез в одних тканях (печень) и липолиз в других (жировое депо). В результате у лиц, получающих ГКГ, в крови возрастает уровень ВЖК, триглицеридов, что стимулирует гиперлипидемию и кетоз.
Влияние ГКГ на обмен белков и нуклеиновых кислот
В печени ГКГ в целом оказывают анаболическое действие на обмен белков и нуклеиновых кислот, т.е. усиливают их биосинтез, а в других тканях они вызывают противоположный – катаболический эффект.
11-оксигенированные ГКГ усиливают секрецию соляной кислоты и пепсиногена слизистой желудка и трипсиногена поджелудочной железой. Эти эффекты ГКГ могут стать причиной язвенных повреждений желудка и кишечного тракта уже через несколько дней стероидной терапии, обильного кровотечения и гибели пациента. Так погиб пациент с катарактой (практически здоровый) от профузного кровотечения в желудочно-кишечном тракте на 3-й день гормонотерапии, назначенной с целью ускорения созревания катаракты.
Влияние минералокортикоидных гормонов (альдостерона) на обмен электролитов и воды
Минералокортикоиды стимулируют обратный транспорт (реабсорбцию) натрия, хлора и НСО3 в дистальных извитых канальцах и собирательных трубочках почек, слюнных, потовых железах, слизистой желудочно-кишечного тракта. Он также способствует выделению (секреции) почками K+, H+, NH4+. Активность альдостерона в этом плане превышает активность 11-дезоксикортикостерона в 30-50 раз, а кортизола и кортикостерона даже в 1000 раз. Альдостерон - самый мощный из природных минералокортикоидов. Кортизол менее активен, однако врачу нельзя забывать, что его вырабатывается гораздо больше, и он тоже оказывает существенное влияние на обмен минеральных веществ.
Предполагается следующий механизм обратного транспорта натрия и влияния на него, прежде всего альдостерона:
1) из первичной мочи, омывающей апикальную поверхность почечных клеток, Na переходит пассивно. Альдостерон лишь повышает число натриевых каналов на апикальной стороне мембран этих клеток;
2) далее происходит перенос иона (Na) из клетки в интерстициальную жидкость с помощью Na/К-зависимой АТФ-азы, синтез которой усиливается по схеме: гормон → рецептор в клетке → хроматин → м-РНК → усиление синтеза белка-энзима Na/К-АТФ-азы.
I. Избыточное введение любого из кортикостероидов, и особенно альдостерона, сопровождается повышением содержания внеклеточного натрия, а с ним увеличение объема внеклеточной жидкости. Одновременно это ведет к уменьшению калия в сыворотке крови. Более того, калий внутри клеток заменяется на натрий. В результате возрастает кровяное давление, и это оказывает неблагоприятное действие на сердечную мышцу.
II. При недостаточной функции коры надпочечников и снижении поступления в кровь названных гормонов понижается канальцевая реабсорбция натрия – это ведет к увеличению экскреции натрия, хлора, ↓ воды и уменьшению ↓ объема (гиповолемии) плазмы. Сгущение крови, повышение ее вязкости сопровождается уменьшением эффективности работы сердца. Перераспределение ионов в сыворотке и тканях ведет к гиповолемии. В связи с потерей жидкости нарушается кровоснабжение почек и развивается почечная недостаточность. Концентрация мочевины в крови больных от этого повышается.
Врач, назначая пациенту гормоны коры надпочечников, обязан контролировать их метаболические эффекты действия и во-время изменять дозу, время и наименование гормона или отменять их.
НАРУШЕНИЯ ФУНКЦИИ КОРЫ НАДПОЧЕЧНИКОВ И ЗАБОЛЕВАНИЯ
I. - Первичная недостаточность, т.е. гипофункция коры надпочечников ведет к развитию Аддисоновой болезни. Она сопровождается потерей натрия с мочой и гипогликемией. Причиной последнего является крайне высокая чувствительность к инсулину вследствие недостаточного содержания глюкокортикоидов. Больные жалуются на анорексию, потерю веса, тошноту, резко выраженные слабость и непереносимость стресса. Понижаются АД, скорость клубочковой фильтрации и способность справиться с нагрузкой водой, тяга к соленому, т.к. уровень натрия в плазме понижен, а калия увеличен. У них повышена пигментация кожи и слизистых, что обусловлено компенсаторно повышенной секрецией АКТГ и ПОМК (отсюда второе название – Бронзовая болезнь). Диета с большим количеством натрия и незначительным содержанием калия компенсирует дисбаланс содержания электролитов и позволяет предотвратить неизбежный летальный исход.
II. Вторичная недостаточность надпочечников обусловлена дефицитом АКТГ при опухоли, инфаркте или инфекции вгипофизе. У пациентов те же симптомы, что и при первичной недостаточности, но отсутствует гиперпигментация.
III. - Избыток глюкортикоидов сопровождается синдромом Кушинга Чаще всего это является следствием фармакологического использования стероидов, а также усиленной секреции АКТГ при аденоме гипофиза или карциноме надпочечников.
Гормоны мозгового слоя надпочечников (катехоламины - адреналин и норадреналин).
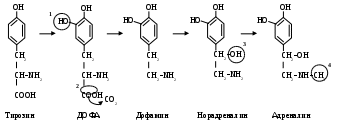
Биосинтез катехоламинов из тирозина происходит в хромафинных клетках мозгового слоя надпочечников. Главный продукт – адреналин (
80%). Вне мозгового слоя надпочечников адреналин не образуется. В противоположность к нему основное количество норадреналина образуется в нервных окончаниях (синапсах).
Превращение тирозина в адреналин включает 4 реакции: (особенности обмена тирозина)
гидроксилирование кольца тирозина с образованием ДОФА;
декарбоксилирование ДОФА с образованием дофамина;
гидроксилирование боковой цепи дофамина – образуется при этом норадреналин;
N- метилирование норадреналина – образуется адреналин.
Е1
Тирозин-гидроксилаза лимитирует скорость всего процесса биосинтеза катехоламинов.
Регулируют образование катехоламинов также клетки чревного нерва, волокна которого иннервируют мозговой слой надпочечников. Синтезированные катехоламины накапливаются в хромаффинных гранулах. Нервная стимуляция мозгового слоя надпочечников приводит к выбросу их из гранул в русло крови путем экзоцитоза. Поэтому при стрессе их концентрация может очень быстро повышаться на несколько порядков (в 1000 раз).
В крови катехоламины циркулируют в слабо диссоциированном виде с альбуминами. Их период полужизни составляет всего 10-30 секунд. Катехоламины не проникают через гематоэнцефалический барьер, и, следовательно, их присутствие в ЦНС – следствие местного синтеза. При болезни Паркинсона нарушается синтез дофамина в мозге. Предшественник его ДОФА – легко преодолевает гематоэнцефалический барьер и поэтому служит эффективным средством лечения болезни Паркинсона.
Действуют катехоламины через два класса рецепторов:
α -адренергические (подклассы α1 и α2 рецепторы)
β- адренергические (подклассы β1 и β2)
Катехоламины, связываясь с рецепторами α2 ингибируют аденилатциклазу и образование ц-АМФ, а, связываясь с β1 и β2 – стимулируют этот энзим и усиливают образование ц-АМФ. По этой причине эффект их действия различен:
α1
Вторичные мессенд-жеры: Са, фосфа-тидилинозит
|
α2
|
β1
|
β2
|
Вторичный мессенджер – ц АМФ
|
Ингибирует аденилатциклазу
|
Активирует аденилатциклазу
|
-1) повышение гликогенолиза (мобилизация гликогена)
- 2) сокращение гладких мышц кровеносных сосудов и мочеполовой системы
- гликоген → глюкагон
|
-1)ингибирование липолиза
- 2) расслабление гладких мышц желудочно-кишечного тракта
Липолиз →
|
- 1)стимуляция липолиза
-2) увеличение силы и амплитуды сокращений миокарда
Липолиз → Тг
|
-1) повышение гликогенолиза в печени и склетных мышцах
- 2)стимуляция глюконеогенеза
- увеличение секреции инсулина, глюкагона, ренина
-3) расслабление гладких мышц бронхов, кровеносных сосудов и желудочно-кишечного тракта
- гликоген – глюкоза
- глюконеогенез
| |
|
|
 Скачать 0.67 Mb.
Скачать 0.67 Mb.