6 итог по патшизе. Вариант 4 Виды расстройств функции эндокринных желез, их характеристика. Типы гормональных эффектов, их харка
 Скачать 6.14 Mb. Скачать 6.14 Mb.
|

|
Структурные компоненты ноцицептивной системы и их характеристика. Структурно-функциональная организация ноцицептивной системы 1. Рецепторный аппарат боли. 2. Проводниковый аппарат ноцицептивной системы. 3. Центральный аппарат системы боли. 4. Медиаторы боли. Виды болевых рецепторов I. По характеру активации: 1) Мономодальные механические; 2) Бимодальные механические и термические; 3) Полимодальные. II. По механизму возбуждения: 1) Механорецепторы; 2) Хемомеханорецепторы. «Молчащие» рецепторы. Виды альгогенов по происхождению 1) тканевые альгогены (гистамин); 2) альгогены плазмы крови (брадикинин); 3) альгогены, выделяющиеся из периферических окончаний С-ноцицепторов (субстанция P). Проводники болевых импульсов Волокна нейронов спинальных ганглиев: 1) Тонкие миелинизированные А-дельта волокна – диаметр 1-4 мкм, скорость проведения воздуждения 5-30 м/с. 2) Безмиелиновые С-волокна –диаметр менее 1 мкм, скорость проведения возбуждения – 0,5-2 м/с. Нейроны болевой чувствительности задних рогов спинного мозга: 1) Специфические ноцицептивные нейроны. 2) Нейроны широкого динамического диапазона. 3) Интернейроны желатинозной субстанции. Медиаторы боли: 1) возбуждающие аминокислоты (глутамат, аспартат); 2) субстанция Р; 3) нейрокинин А; 4) CGRP (calcitonin gene-related peptide); 5) оксид азота. Пути проведения болевых импульсов в ЦНС: 1) Неоспиноталамический тракт (латеральный); 2)Палеоспиноталамический тракт (медиальный, спиноретикулоталамиче-ский) 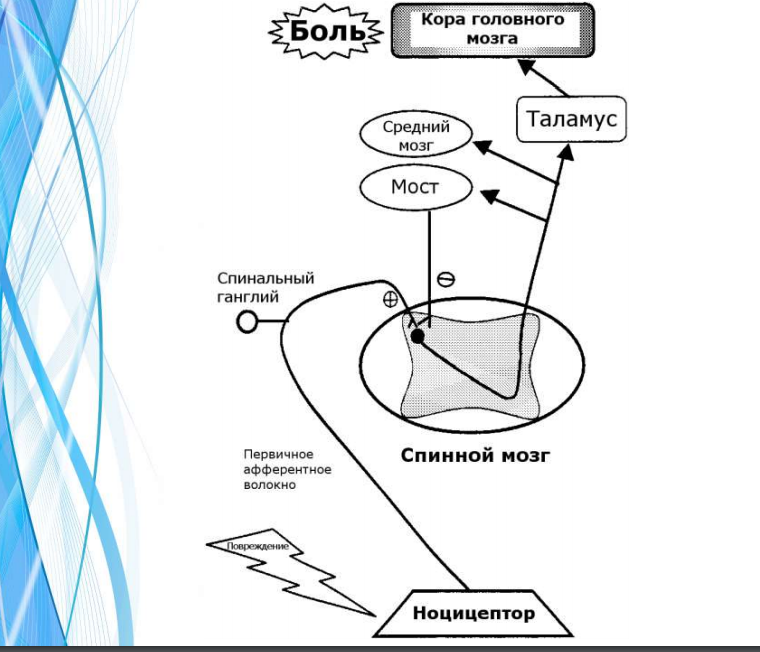 6 вариант 1.роль механизмов обратной связи в саморегуляции диятельности эндокринных желез. Типы механизмов обратной связи Наряду с факторами прямой регуляции функций желез внутренней секреции важную роль в их деятельности играют механизмы саморегуляции - механизмы обратной связи в регуляции функции желез . Так, гонадотропины гипофиза специфически усиливают эндокринную функцию яичников и, в частности, секрецию ими эстронов (плюс-действие), высокие же концентрации эстрогенов в крови тормозят секрецию гипофизом гонадотропинов (минус-действие). Прямая регуляция эндокринной железы может быть не только положительной, но и отрицательной, ингибиторной. Примером тормозного влияния могут служить эффектысоматостатина гипоталамуса на секрецию СТГ , гипоталамического лактостатина на секрециюпролактина , глюкозы на секрецию глюкагона поджелудочной железы или ионов кальция на секрецию паратгормона околощитовидными железами . В этих случаях снижение секреции гормонов тормозит действие отрицательных регуляторных стимулов и таким образом нормализует функционирование железы. Обратная связь при некотором уровне концентрации гормонов может быть не только отрицательной, но и положительной ("плюс-плюс взаимодействие"). В данном случае регуляция и саморегуляция способствуют усилению работы функциональной системы, ее самоактивации. Клинические синдромы, развивающиеся при гиперфункции мозгового слоя коры надпочечников. Возникает чаще всего при опухоли хромаффинной ткани – феохромоцитоме. В опухоли обычно образуется избыточное количество адреналина и норадреналина, которые и обусловливают клинику. При этом выделяют ряд синдромов: сердечно-сосудистый (изменение деятельности сердца, тонуса сосудов); нервно-психический (галлюцинации, головные боли, головокружение, повышенная возбудимость и т.д.); желудочно-кишечный (тошнота, рвота, запоры, язвенные явления с кровотечениями); нарушения обмена веществ. Болезнь протекает в виде пароксизмов. 3.Денервационный и диафферантационный синдромы. Механизмы и проявления А)Денервационный синдром представляет собой комплекс изменений, возникающих в постсинаптических нейронах, органах и тканях после выпадения нервных влияний на эти структуры. Денервированная структура (мышца, нейрон) приобретает повышенную чувствительность к физиологически активным веществам (закон Кеннона-Розенблюта). Основным проявлением денервационного синдрома в мышце является исчезновение концевой пластинки - зоны мышечного волокна, где сосредоточен весь его холинергический аппарат. Вместо нее появляются новые ацетилхолиновые рецепторы на всем протяжении мышечного волокна, и в связи с этим происходит повышение общей чувствительности к ацетилхолину всего волокна (А.Г. Гинецинский, Н.М. Ашмарина). Этот эффект связан главным образом с выпадением трофических влияний нерва. Другой характерный признак - фибриллярные подергивания денервированной мышцы. Этот эффект отражает реакцию мышечных денервированных волокон на поступающий к ним из разных сторонних источников ацетилхолин.
При денервации в мышце и других тканях появляются свойства, присущие ранним, в частности, эмбриональным стадиям развития. Это явление возникает как результат патологического растормаживания супрессированных в норме генов. Б)Импульсация, поступающая в нейрон, из какого бы источника она ни исходила, является для нейрона афферентной. Выключение этой афферентации представляет собой деафферентацию нейрона. Последняя может быть обусловлена либо выпадением поступающей импульсации (при перерыве нервных путей, нарушении выделения нейромедиаторов пресинаптическими окончаниями), либо блокадой воспринимающих рецепторов на постсинаптическом нейроне (при действии токсинов, фармакологических средств и др.). Многие явления при деафферентации нейрона представляют собой выражение денервационного синдрома. Полной деафферентации нейрона не происходит, так как нейроны ЦНС обладают огромным количеством афферентных входов. Тем не менее и при частичной деафферентации возникает повышение возбудимости нейрона или его отдельных участков и нарушение тормозных механизмов. В силу этого при деафферентации группа нейронов может приобрести свойства ГПУВ. В клинике под феноменом деафферентации имеют в виду синдромы, возникающие при выпадении афферентной стимуляции с периферии. Эти синдромы можно воспроизвести в эксперименте путем перерезки соответствующих задних корешков спинного мозга. Движения конечности, иннервируемой деафферентированными таким образом сегментами спинного мозга, становятся размашистыми, плохо координированными. Кроме того, такая конечность способна осуществлять спонтанные движения в такт с дыханием (феномен Орбели-Кунстман), что обусловлено растормаживанием и повышением возбудимости деафферентированных нейронов спинного мозга. 4.Виды боли различные классификации. Свойства эпикритической и протопатической боли. Классификация боли: I. По происхождению: А) «Физиологическая» - вызванная определённым внешним воздействием; - зависит от силы и характера раздражителя (адекватна ему); - мобилизует защитные силы организма; - является сигналом опасности (возможность повреждения). Б) Патологическая = нейропатическая - вызванная повреждением нерв. системы; - не адекватна определённому воздействию; - не мобилизует защитные силы организма - является сигналом патологии, характерна для заболеваний нервной системы. II. По локализации ноцицепторов и характеру болевых ощущений: 1. Соматическая: а) поверхностная: - эпикритическая (ранняя, быстрая); - протопатическая (поздняя, медленная). б) глубокая. 2. Висцеральная: (связана с зонами Захарьина-Геда) а) истинная; б) отраженная. Соматическая боль связана с повреждениями кожи, мышц, ОДА в целом. Поверхностная боль возникает при раздражении ноцицепторов кожи, Эпикритическая (ранняя) боль называется быстрой потому, что: возникает через доли секунды; имеет короткий латентный период; точно локализуется; быстро проходит; острое быстрое преходящее ощущение. Протопатическая (поздняя) боль характеризуется: более продолжительным латентным периодом (несколько секунд); более диффузная; более продолжительная; сопровождается неприятным болевым ощущением. Такое разделение связано с проведением возбуждения – по миелиновым волокнам А  (быстрая боль); по безмиелиновым волокнам С (медленная боль). (быстрая боль); по безмиелиновым волокнам С (медленная боль).Волокна группы А – толстые миелиновые волокна (Vпров. 50-140 м/сек). Волокна группы В – меньшего диаметра, В1 и В2 (Vпров. 15-30; 10-15 м/сек). Волокна С – безмиелиновые – меньшего диаметра (V=0,6-2 м/сек). Безмиелиновые волокна более устойчивы: - к гипоксии (т.к. снижена активность метаболизма); - быстрее регенерируют; - характеризуются более диффузным распределением волокон в зоне иннервации. При сдавлении нервных волокон в первую очередь страдают миелиновые волокна, анестетик при проведении анестезии быстрее подействует на безмиелиновые волокна. Глубокая боль связана с раздражением рецепторов глубоких тканей (сухожилия, кости, надкостница). Характер боли: - тупые; - ноющие; - длительные; - диффузные; - склонны к иррадиации. Причины глубокой боли: - растяжение тканей; - сильное давление на ткани; - ишемия; - действие химических раздражителей. Висцеральная боль – возникает при раздражении рецепторов внутренних органов. Характер болей: - тупые; - ноющие; - мучительные; - продолжительные; - высокая способность к иррадиации. Причины висцеральных болей: - растяжение полых органов; - спастические сокращения полых органов; - растяжение (спастическое сокращение кровеносных сосудов органов); - ишемия; - химическое раздражение оболочек органов (при ЯБ); - сильное сокращение органов (сокращение кишечника Вариант 7 Железистые и внежелезистые причины эндокринопатий. Их характеристика "Железистые" причины. Данная группа причин включает факторы, прямо повреждающие ткань железы: опухоли, травмы, воспалительные процессы, расстройства кровообращения, гипоксии, инфекции (в том числе вирусные), интоксикации, склерозирование железы, удаление железы, длительная бездеятельность, истощение, а также наследственные дефекты синтеза гормонов. Периферические (внежелезистые) причины: нарушение инактивации гормонов в тканях и печени; нарушение связывания гормонов с белками; образование антител к гормону; нарушение соединения гормона со своими рецепторами в клетках-мишенях; существование антигормонов и их действие на рецепторы по механизму конкурентного связывания. Антигормоны – вещества, в том числе гормоны, которые имеют сродство к рецепторам данного гормона и взаимодействуют с ними. Занимая рецепторы, они блокируют эффект данного гормона. Примером антигормонов могут служить андрогены и эстрогены. (скопировала из пособия Северьяновой, не знаю что с характеристиками делать) механизмы нарушений белкового, жирового, энергетического и углеводного при гипотиреозе Дефицит гормонов щитовидной железы приводит к нарушению всех видов обмена: белкового (понижение синтеза и распада белка), углеводного (повышенная толерантность к углеводам, склонность к гипогликемии), липидного (увеличение в крови липопротеидов и холестерина), водно-солевого (задержка воды и натрия в тканях. Задержка воды и натрия, накопление в соединительной ткани мукопротеидов с выраженными гидрофильными свойствами приводят к развитию слизистого отека. В основе всех нарушений лежит снижение окислительных процессов и энергетическая недостаточность, обуславливающая понижение синтеза белков и прекращение роста и физического развития. Нарушение функции вегетативной нервной системы (угнетение ее симпатического отдела) приводит к функциональным сдвигам в деятельности многих органов и систем (брадикардия, снижение моторики ЖКТ, уменьшение потоотделения и т.д.). Наблюдается также снижение психической активности и интеллекта – гипотиреоидная энцефалопатия. Тяжелым осложнением является развитие гипотиреоидной комы, для которой характерно угнетение дыхательного центра, прогрессирующее снижение сердечного выброса, гипоксия мозга и гипотермия, полисерозит (накопление жидкости в серозных полостях). (северьянова)  трофогены: понятие, источники, основные виды трофических веществ. Сущность представлений о нормо и патотрофогенах трофогены – это вещества различной, преимущественно белковой природы, осуществляющие собственно трофические эффекты поддержания жизнедеятельности и генетически заложенных свойств клетки источники трофогенов: нейроны, периферические клетки, глиальные клетки, шванновские клетки. Вещества, играющие роль трофогенов, образуются также из сывороточных и иммунных белков. Трофическое воздействие могут оказывать некоторые гормоны. В регуляции трофических процессов принимают участие пептиды, ганглиозиды, некоторые нейромедиаторы.   сущность представлений о воротной регуляции болевого потока на входе в спинной мозг 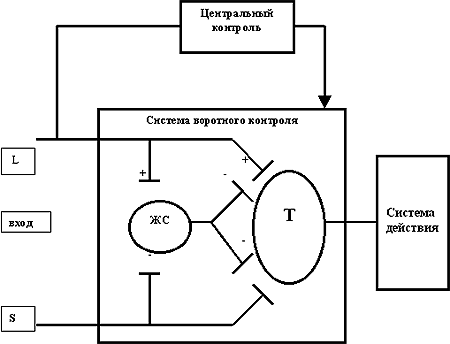 ЖС - желатинозная субстанция задних рогов спинного мозга Т - трансмиссивные нейроны L - волокна большого диаметра S - волокна малого диаметра Рис. 1. Схема воротного механизма контроля боли (R.Melzack, 1999) 1.Передача нервных импульсов в центральную нервную систему модулируется специальными "воротными" механизмами, расположенными в задних рогах спинного мозга. 2.Спинальные воротные механизмы представляют собой взаимосвязь активности афферентных волокон большого диаметра (L) и волокон малого диаметра (S): активность L-волокон тормозит передачу импульсов ("закрывает ворота"), в то время как активность S-волокон облегчает их передачу ("открывает ворота"). 3.Спинальные "воротные" механизмы, в свою очередь, также регулируются нисходящими импульсами от головного мозга, активируемыми системой быстропроводящих волокон большого диаметра (L). 4.При достижении критического уровня поток импульсов от нейронов спинного мозга (релейные, трансмиссивные нейроны, передаточные Т-клетки) активирует Систему действия, т.е. те нейрональные зоны центральной нервной системы, которые формируют сложные поведенческие реакции на боль. Вариант 8 Процессы, составляющие эндокринную функцию. Генетические обусловленные дефекты биосинтеза гормона I. Биосинтез гормонов. Известны два основных типа генетического контроля синтеза гормонов: 1. Прямой контроль – синтез в полисомах предшественников белково-пептидных гормонов, характеризующийся схемой: ген → мРНК → прогормон → гормон. 2. Опосредованный контроль по схеме: гены → мРНК → фермент → гормон. II. Секреция. Секреция многих гормонов осуществляется импульсно, дискретными порциями. При этом секреторный процесс может включать различные механизмы: 1. Освобождение гормона из секреторных гранул, происходящее с использованием энергии АТФ и при участии ионов кальция (белково-пептидные гормоны, катехоламины). 2. Освобождение из белковосвязанной формы (тиреоидные гормоны).3. Экзоцитоз по схеме: миграция гранул, содержащих гормон, к мембране клетки → растворение мембраны в месте контакта → прорыв содержимого гранулы в межклеточное пространство → диффузия в капилляры.4. Диффузия через плазматические мембраны (стероидные гормоны). III. Транспорт гормонов кровью. Происходит в двух формах: свободной и связанной. В отношении транспорта в связанной форме выделяют: 1.Специфическое связывание со специальными транспортными белками 2.Неспецифическое связывание – связывание с гамма-глобулинами, ад-сорбция на эритроцитах. Биологическое значение связывания: предотвращение чрезмерного гормонального эффекта; создание биологического резерва гормона; защита гормона от быстрой инактивации; защита гормона от быстрого выведения. IV. Взаимодействие гормона с клеткой-мишенью включает процесс рецепции и трансдукции. Рецепция основана на существовании на поверхности или внутри клетки специальных веществ, обладающих высокой избирательной чувствительностью (аффинностью) к сигнальной молекуле-лиганду. В зависимости от локализации рецепторов различают два основных типа взаимодействия: мембранный (контактный, внеклеточный) и внутриклеточ-ный, включающий два варианта: цитозольный и ядерный . 1. Мембранный тип рецепции характерен для белковых и полипептидных гормонов, для которых мембрана клеток непроницаема. Последовательность взаимодействий при этом: взаимодействие гормона и рецептора → образование активного комплекса "гормон + рецептор" → активация соответствующего фермента, включающего определенную систему вторичных (вторых) посредников, которые обеспечивают формирование ответа клетки на сигнал (гормон). 2. Внутриклеточный тип рецепции. А. Цитозольный тип внутриклеточной рецепции. Характерен для стероидных гормонов, проникающих через мембрану клеток. Схема взаимодействия: образование активного комплекса "гормон + рецептор" → транслокация комплекса в ядро клетки и связывание с хроматиновым рецептором → избирательная инициация транскрипции мРНК → изменение белкового синтеза. Б. Ядерный тип внутриклеточной рецепции установлен для тиреоидных гормонов. Внутриклеточные рецепторы локализованы в ядре клетки и связаны с хроматином. Связывание рецепторов с тиреоидными гормонами усиливает экспрессию генетической информации с усилением синтеза специфической мРНК, кодирующей ферментные белки соответствующего биологического ответа. Трансдукция. Образование комплекса гормон – рецептор инициирует активацию внутриклеточных процессов, обеспечивающих в конечном итоге ответ клетки, обусловленный ее спецификой. Это явление получило название трансдукции биологического сигнала в клеточный ответ. V. Инактивация гормонов. В этом конечном этапе любой эндокринной функции принимают участие обменные процессы, протекающие при использовании гормона в реагирующих тканях (активация, реактивация, взаимопревращения), т.е. периферические превращения и катаболизм, а также специальные процессы инактивации. Генетически обусловленные дефекты биосинтеза гомонов: 1. Гипофункция железы 2. Накопление в железе образующихся до места блокады промежуточных продуктов биосинтеза 3. Нарушение механизма обратной связи и развитие дополнительных патологических процессов. |