Основные системы межклеточной коммуникации: эндокринная, паракринная, аутокринная системы.
Для нормального функционирования многоклеточного организма необходима взаимосвязь между отдельными клетками, тканями и органами. Эту взаимосвязь осуществляют 4 основные системы регуляции (рис. 11-1). Центральная и периферическая нервные системы через нервные импульсы и нейромедиаторы; Эндокринная система через эндокринные железы и гормоны, которые секретируются в кровь и влияют на метаболизм различных клеток-мишеней; Паракринная и аутокринная системы посредством различных соединений, которые секретируются в межклеточное пространство и взаимодействуют с рецепторами либо близлежащих клеток, либо той же клетки (простагландины, гормоны ЖКТ, гистамин и др.); Иммунная система через специфические белки (цитокины, антитела).
Нервная и гуморальная регуляция как единая система регуляции обмена веществ. Гормоны – первичные посредники в передаче информации.
Механизмы регуляции физиологических функций традиционно подразделяют на нервные и гуморальные, хотя в действительности они образуют единую регуляторную систему, обеспечивающую поддержание гомеостаза и приспособительную деятельность организма. Эти механизмы имеют многочисленные связи как на уровне функционирования нервных центров, так и при передаче сигнальной информации эффекторным структурам. Достаточно сказать, что при осуществлении простейшего рефлекса как элементарного механизма нервных регуляций передача сигнализации с одной клетки на другую осуществляется посредством гуморальных факторов - нейромедиаторов. Чувствительность сенсорных рецепторов к действию раздражителей и функциональное состояние нейронов изменяется под действием гормонов, нейромедиаторов, ряда других биологически активных веществ, а также простейших метаболитов и минеральных ионов (К+, Na+, Ca-+, С1). В свою очередь, нервная система может запускать или выполнять коррекцию гуморальных регуляций. Гуморальные регуляции в организме находятся под контролем нервной системы. Гуморальные механизмы филогенетически более древние, они имеются даже у одноклеточных животных и приобретают большое разнообразие у многоклеточных и особенно у человека. Нервные механизмы регуляций образовались филогенетически и формируются постепенно в онтогенезе человека. Такие регуляции возможны лишь в многоклеточных структурах, имеющих нервные клетки, объединяющиеся в нервные цепи и составляющие рефлекторные дуги. Гуморальные регуляции осуществляются путем распространения сигнальных молекул в жидкостях организма по принципу "всем, всем, всем", или принципу "радиосвязи". Нервные регуляции осуществляются по принципу "письмо с адресом", или "телеграфной связи". Сигнализация передается от нервных центров к строго определенным структурам, например к точно определенным мышечным волокнам или их группам в конкретной мышце. Только в этом случае возможны целенаправленные, координированные движения человека. Гуморальные регуляции, как правило, осуществляются медленнее, чем нервные. Скорость проведения сигнала (потенциала действия) в быстрых нервных волокнах достигает 120 м/с, в то время как скорость транспорта сигнальной молекулы с током крови в артериях приблизительно в 200 раз, а в капиллярах - в тысячи раз меньше. Приход нервного импульса к органу-эффектору практически мгновенно вызывает физиологический эффект (например, сокращение скелетной мышцы). Реакция на многие гормональные сигналы более медленная. Например, проявление ответной реакции на действие гормонов щитовидной железы и коры надпочечников происходит через десятки минут и даже часы. Гуморальные механизмы имеют преимущественное значение в регуляции процессов обмена веществ, скорости деления клеток, роста и специализации тканей, полового созревания, адаптации к изменению условий внешней среды. Нервная система в здоровом организме оказывает влияние на все гуморальные регуляции, осуществляет их коррекцию. Вместе с тем у нервной системы имеются свои специфические функции. Она регулирует жизненные процессы, требующие быстрых реакций, обеспечивает восприятие сигналов, приходящих от сенсорных рецепторов органов чувств, кожи и внутренних органов. Регулирует тонус и сокращения скелетных мышц, которые обеспечивают поддержание позы и перемещение тела в пространстве. Нервная система обеспечивает проявление таких психических функций, как ощущение, эмоции, мотивации, память, мышление, сознание, регулирует поведенческие реакции, направленные на достижение полезного
приспособительного результата. Безруких М.М. Возрастная физиология: Учебное пособие для вузов.- М.: ACADEMIA, 2003.- С. 44.
Интегрирующими регуляторами, связывающими различные регуляторные механизмы и метаболизм в разных органах, являются гормоны. Они функционируют как химические посредники, переносящие сигналы, возникающие в различных органах и ЦНС. Ответная реакция клетки на действие гормона очень разнообразна и определяется как химическим строением гормона, так и типом клетки, на которую направлено действие гормона.
В крови гормоны присутствуют в очень низкой концентрации. Для того чтобы передавать сигналы в клетки, гормоны должны распознаваться и связываться особыми белками клетки - рецепторами, обладающими высокой специфичностью.
Физиологический эффект гормона определяется разными факторами, например концентрацией гормона (которая определяется скоростью инактивации в результате распада гормонов, протекающего в основном в печени, и скоростью выведения гормонов и его метаболитов из организма), его сродством к белкам- переносчикам (стероидные и тиреоидные гормоны транспортируются по кровеносному руслу В комплексе с белками), количеством и типом рецепторов на поверхности клеток-мишеней.
Регуляция синтеза и секреции гормонов по принципу обратной связи.
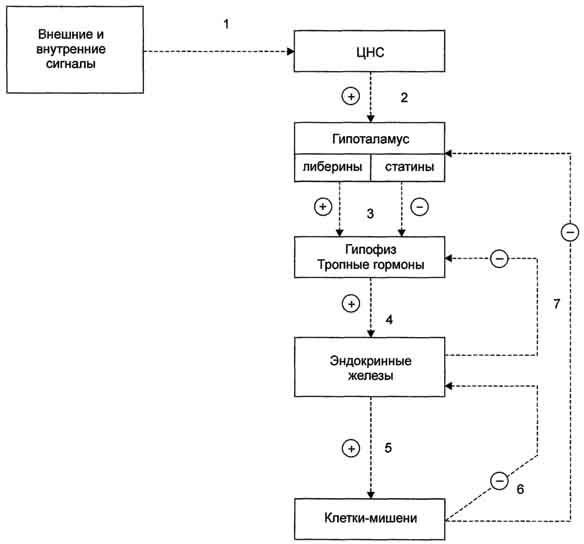
Отечественный ученый М.М. Завадовский, изучая закономерности в регуляции деятельности эндокринных желез, впервые в 1933 г. сформулировал принцип “плюс-минус взаимодействие”, получивший в дальнейшем название “принцип обратной связи”.
Под обратной связью подразумевается система, в которой конечный продукт деятельности этой системы (например, гормон, нейротрансмиттер и другие вещества) модифицирует или видоизменяет функцию компонентов, составляющих систему, направленную на изменение количества конечного продукта (гормона) или активности системы. Жизнедеятельность всего организма является следствием функционирования многочисленных саморегулируемых систем (выделительная, сердечно-сосудистая, пищеварительная, дыхательная и др.), находящихся в свою очередь под контролем нейроэндокринно-иммунной системы. Все перечисленное представляет, таким образом, комплекс различных саморегулируемых систем, находящихся в
определенной степени зависимости и “подчиненности”. Конечный результат или активность системы может модифицироваться двумя путями, а именно путем стимуляции для увеличения количества конечного продукта (гормона) или повышения активности эффекта, или путем угнетения (ингибирования) системы с целью уменьшения количества конечного продукта или активности. Первый путь модифицирования называется положительной, а второй – отрицательной обратной связью. Примером положительной обратной связи является повышение уровня гормона в крови, стимулирующее высвобождение другого гормона (повышение уровня эстрадиола в крови вызывает высвобождение ЛГ в гипофизе), а отрицательной обратной связи, когда повышенный уровень одного гормона угнетает секрецию и высвобождение другого (повышение концентрации тироидных гормонов в крови снижает секрецию ТТГ в гипофизе).
Клетки-мишени и клеточные рецепторы гормонов. Рецепторы цитоплазматической мембраны. Рецепторы, локализованные в цитоплазме. Рецепторы сопряженные с ионными каналами. Регуляция работы рецепторного аппарата.
Биологическое действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней. Для проявления биологической активности связывание гормона с рецептором должно приводить к образованию химического сигнала внутри клетки, который вызывает специфический биологический ответ, например изменение скорости синтеза ферментов и других белков или изменение их активности (см. раздел 5). Мишенью для гормона могут служить клетки одной или нескольких тканей. Воздействуя на клетку-мишень, гормон вызывает специфическую ответную реакцию. Например, щитовидная железа - специфическая мишень для тиреотропина, под действием которого увеличивается количество ацинарных клеток щитовидной железы, повышается скорость биосинтеза тиреоидных гормонов. Глюкагон, воздействуя на адипоциты, активирует липолиз, в печени стимулирует мобилизацию гликогена и глюконеогенез. Характерный признак клетки- мишени - способность воспринимать информацию, закодированную в химической структуре гормона.
Начальный этап в действии гормона на клетку-мишень - взаимодействие гормона с рецептором клетки. Концентрация гормонов во внеклеточной жидкости очень низка и обычно колеблется в пределах 10-6-10-11 ммоль/л. Клетки-мишени отличают соответствующий гормон от множества других молекул и гормонов благодаря наличию на клетке-мишени соответствующего рецептора со специфическим центром связывания с гормоном.
Передача гормональных сигналов через мембранные рецепторы
Гормоны (первичные посредники), связываясь с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрации особых молекул внутри клетки - вторичных посредников. Вторичными посредниками могут быть следующие молекулы: цАМФ, цГМФ, ИФ3, ДАТ, Са2+, NO.
Гормоны, взаимодействие которых с рецептором клетки-мишени приводит к образованию цАМФ, действуют через трёхкомпонентную систему, которая включает белок-рецептор, G-белок и фермент аденилатциклазу. Образующийся под действием аденилатциклазы цАМФ активирует протеинкиназу А, фосфорилируюшую ферменты и другие белки (см. раздел 5). Известно более 200 различных G-белков, в структуре которых обнаружены 3 субъединицы α, β и γ (см. раздел 5). В отсутствие гормона α-субъединица G-белка связана с ГДФ. Образование комплекса гормонрецептора приводит к конформационным изменениям α-субъединицы, замене ГДФ на ГТФ и отщеплению димера βγ от α-ГТФ. В случае рецепторов, сопряжённых с Gs-белком, субъединица αs-ГТФ активирует аденилатциклазу (рис. 11-3).
В случае рецепторов, сопряжённых с Giбелком, субъединица αi-ГТФ ингибирует аденилатциклазу. В таблице 11-4 приведены примеры гормонов, взаимодействие которых с соответствующим рецептором активирует или ингибирует аденилатциклазу.
Другая система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Цитоплазматический домен такого типа рецепторов обладает активностью гуанилатциклазы, которая катализирует реакцию образования цГМФ из ГТФ> (подобно аденилатциклазе). Молекулы цГМФ могут активировать ионные каналы либо активировать цГМФ-за-висимую протеинкиназу G, участвующую в фосфорилировании других белков в клетке. Например, фосфодиэстерааа, которая гидролизует цАМФ до АМФ, активируется в результате фосфорилирования цГМФ-зависимой протеинкиназой.
Некоторые гормоны (например, вазопрессин или адреналин), образуя комплекс с соответствующими рецепторами (рецептор V1 для вазопрессина и αi-рецептор для адреналина), через активацию соответствующих G-белков активируют фосфолипазу С, в результате чего в клетке появляются вторичные
посредники ИФ3, ДАГ. Молекула ИФ3 стимулирует высвобождение Са2+ из ЭР. Кальций связывается с белком кальмодулином. Этот комплекс активирует Са2+-кальмодулинзависимую протеинкиназу. Ионы кальция и ДАГ участвуют в активации протеинкиназы С (см. раздел 5).
Многие гормоны передают сигнал в клетку через рецепторы, которые либо обладают тиро-зинкиназной активностью, либо связываются с цитоплазматическими белками, проявляющими активность тирозинкиназы. Связывание инсулина с мембранным рецептором, который является тирозинкиназой и имеет центр фосфорилирования, инициирует аутофосфорилирование и последующее фосфорилирование субстратов рецептора инсулина и других белков (см. разделы 5 и ниже подраздел III, Ж).
В случае взаимодействия, например, эпидермального фактора роста или инсулиноподобного фактора роста - 1с мембранным рецептором сначала происходят димеризация рецептора и его активация. Активированный таким образом гомодимер рецептора, участок которого на внутренней стороне мембраны обладает активностью тирозинкиназы, фосфорилируется сам (аутофосфорилирование) и вызывает фосфорилирование других белков и ферментов, которые участвуют в активации факторов транскрипции генов.
Некоторые гормоны (например, гормон роста, пролактин, интерферон, цитокины) взаимодействуют с мембранными рецепторами, ассоциированными с цитоплазматическими протеинкиназами (так называемыми "Янус-киназами", или киназами семейства JAK). Присоединение гормона вызьшает димеризацию рецептора, присоединение Янус-киназ, их аутофосфорилирование и активацию. Янус-киназы, в свою очередь, фос- форилируют рецептор по остаткам тирозина, в результате чего рецептор связывается с другими белками, например, особыми белками - переносчиками сигнала и активаторами транскрипции (ПСАТ, или STAT - от англ, signal transducer and activator of transcription - переносчик сигнала и активатор транскрипции). Далее следует инициируемый тирозинкиназой каскад реакций фосфорилирования. Белки STAT фосфорилируются, образуют димеры, транспортируются в ядро, где, связываясь со специфическими участками ДНК, участвуют в регуляции транскрипции (рис. 11-4).
Сигнальной молекулой в клетке может служить также оксид азота NO, образующийся в организме из аргинина при участии фермента NO-синтазы, присутствующего в нервной ткани, эндотелии сосудов, тромбоцитах и других тканях (см. раздел 9). Молекула NO может быстро диффундировать через мембрану эндотелиальных клеток, где она синтезируется, в соседние клетки. Действие оксида азота кратковременно, так как Т1/2 NO колеблется в пределах 5-10 с. В крови молекула существует примерно 100 мс, поскольку быстро взаимодействует с молекулярным кислородом, образуя нитрит, который далее превращается в нитрат и экскретируется с мочой. В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в активный центр гуанилатциклазы ионом железа (см. раздел 5), способствуя тем самым быстрому образованию цГМФ. Увеличение концентрации цГМФ в клетках гладких мышц вызывает активацию киназ, что в конечном итоге приводит к расслаблению ГМК сосудов и последующему их расширению. Механизм действия оксида азота объясняет использование нитроглицерина в качестве лекарственного препарата для снятия острых болей в сердце, поскольку нитроглицерин - источник образующихся молекул NO, которые и вызывают расслабление кровеносных сосудов и увеличение притока крови в миокард.
Передача сигналов через внутриклеточные рецепторы
Стероидные и тиреоидные гормоны связываются с рецепторами внутри клетки и регулируют скорость транскрипции специфических генов (рис. 11-5).
В отсутствие гормона внутриклеточные рецепторы связаны обычно с другими белками в цитозоле или ядре. Например, рецепторы глюкокортикоидов образуют в цитозоле комплекс с шапероном, что препятствует связыванию рецептора с молекулой ДНК (рис. 11-6).
Взаимодействие гормона с центром связывания на С-концевом участке полипептидной цепи рецептора вызывает конформационные изменения и освобождение рецептора от шаперона. Происходит объединение 2 молекул рецептора с образованием гомодимера. Димер рецептора узнаёт специфическую последовательность нуклеотидов, которая расположена в промоторной области гена. Взаимодействие со специфическим участком ДНК HRE (от англ, hormone response element, элемент, реагирующий на воздействие гормона) обеспечивает центральный домен рецептора. Этот домен содержит аминокислотную последовательность, образующую 2 "цинковых пальца". В каждом "цинковом пальце" атом цинка связан с 4 остатками цистеина (рис. 11-7).
В структуре одного "цинкового пальца" имеется последовательность аминокислот, отвечающая за связывание с ДНК, а второй "цинковый палец" содержит последовательность аминокислот, участвующую в димеризации рецепторов. Взаимодействие комплекса гормон-рецептор с определённой последовательностью нуклеотидрв в промоторной части ДНК приводит к активации транскрипции.
Рецепторы тиреоидных гормонов всегда связаны с ДНК. В отсутствие гормонов соответствующие рецепторы ингибируют экспрессию генов. Напротив, взаимодействие с гормоном превращает их в активаторы транскрипции.
Передача сигналов через рецепторы, сопряжённые с ионными каналами
Рецепторы, сопряжённые с ионными каналами, являются интегральными мембранными белками, состоящими из нескольких субъединиц. Они действуют одновременно как ионные каналы и как рецепторы, которые способны специфически связывать с внешней стороны эффектор, изменяющий их ионную проводимость. Эффекторами такого типа могут быть гормоны и нейромедиаторы (см. рис. 11-3).
Известны рецепторы для ряда гормонов, ассоциированных с ионными каналами, и большинства медиаторов, среди которых наиболее изучен рецептор ацетилхолина. Рецептор ацетилхолина состоит из пяти цилиндрообразных субъединиц, расположенных в мембране параллельно друг другу:α2, β, γ, δ. Между ними вдоль оси цилиндров находится заполненный молекулами воды канал. Каждая субъединица рецептора состоит из большого количества гидрофобных аминокислотных остатков. Кроме этого, все субъединицы содержат один спирализованный трансмембранный фрагмент, аминокислотные радикалы которого (полярные незаряженные аминокислотные остатки, в основном серии и треонин) выстилают центральный канал рецептора изнутри. В средней части субъединиц, обращённой к каналу, локализованы остатки лейцина. В присутствии ацетилхолина боковые взаимодействия между субъединицами поддерживают канал в открытом состоянии и создают возможность для транспорта ионов. В отсутствие ацетилхолина в результате изменения ориентации субъединиц относительно друг друга канал закрывается, так как выступающие внутрь канала остатки лейцина образуют плотное гидрофобное кольцо, блокируя движение гидратированных ионов в этой области (рис. 11-8).
2. Регуляция количества и активности рецепторов
Концентрация рецепторов внутри клетки или на её поверхности и их сродство к данному гормону в норме регулируются различными способами, а также могут меняться при заболеваниях или при использовании гормонов или их агонистов в качестве лекарственных средств. Например, при воздействии β-адренергических агонистов на клетки в течение нескольких минут в ответ на новое добавление агониста прекращается активация аденилатциклазы, и биологический ответ исчезает. Такое снижение чувствительности рецептора к гормону (десенситизация) может происходить в результате изменения количества рецепторов по механизму понижающей регуляции. Гормон связывается с рецептором, комплекс гормон-рецептор путём эндоцитоза проникает в клетку (интернализуется), где часть рецепторов подвергается протеолитическому расщеплению под действием ферментов лизосом, а часть инактивируется, отделяясь от других мембранных компонентов. Это приводит к уменьшению количества рецепторов на плазматической мембране. Например, в случае инсулина, глюкагона, катехоламинов это происходит в течение нескольких минут или часов. При снижении концентрации гормона рецепторы возвращаются на поверхность клетки, и чувствительность к гормону восстанавливается. Активность рецептора, т.е. его сродство к гормону, может изменяться также в результате ковалентной модификации, главным образом путём фосфорилирования. Концентрация внутриклеточных рецепторов может также регулироваться по механизму индукции и репрессии.
5. Механизмы передачи гормональных сигналов в клетки: G белки, циклические АМФ и ГМФ как вторичные посредники. Протеинкиназа А и протеинкиназа G.
Механизмы передачи гормональных сигналов в клетки
По механизму действия гормоны можно разделить на 2 группы. К первой группе относят гормоны, взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, а также гормоны местного действия - цитокины, эйкозаноиды). Вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами.
Связывание гормона (первичного посредника) с рецептором приводит к изменению кон-формации рецептора. Это изменение улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путём изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма:
в результате изменения активности ферментов;
в результате изменения количества ферментов (табл. 11-3).
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоящие из α, β и γ-субъединиц. Состав димеров βγ незначительно различаются в разных тканях, но в пределах одной клетки все G-белки, как правило, имеют одинаковый комплект βγ-субъединиц. Поэтому G-белки принято различать по их α-субъединицам. Выявлено 16 генов, кодирующих различные α-субъединицы G-белков. Некоторые из генов имеют более одного белка,, вследствие альтернативного сплайсинга РНК.
Каждая α-субъединица в составе G-белка имеет специфические центры:
связывания ГТФ или ГДФ; взаимодействия с рецептором; связывания с βγ-субъединицами;
фосфорилирования под действием протеинкиназы С;
взаимодействия с ферментом аденилатциклазой или фосфолипазой С.
В структуре G-белков отсутствуют α-спиральные, пронизывающие мембрану домены. G-белки относят к группе "заякоренных" белков (рис. 5-34).
Регуляция активности G-белков
Различают неактивную форму G-белка - комплекс αβγ-ГДФ и активированную форму αβγ-ГТФ. Активация G-белка происходит при взаимодействии с комплексом активатор-рецептор, изменение конформации G- белка снижает сродство α-субъединицы к молекуле ГДФ и увеличивает к ГТФ. Замена ГДФ на ГТФ в активном центре G-белка нарушает комплементарность между α-ГТФ и βγ-субъединицами. Рецептор, связанный с сигнальной молекулой, может активировать большое количество молекул G-белка, таким образом обеспечивая усиление внеклеточного сигнала на этом этапе (рис. 5-35).
Активированная α-субъединица G-белка (α-ГТФ) взаимодействует со специфическим
белком клеточной мембраны и изменяет его активность. Такими белками могут быть ферменты аденилатциклаза, фосфолипаза С, фосфоди-эстераза цГМФ, Nа+-каналы, К+-каналы.
Следующий этап цикла функционирования G-белка - дефосфорилирование ГТФ, связанного с α- субъединицей, причём фермент, катализирующий эту реакцию, - сама α-субъединица.
Дефосфорилирование приводит к образованию комплекса α-ГДФ, который не комплементарен специфическому белку мембраны (например, аденилатциклазе), но имеет высокое сродство к ру-протомерам. G-белок возвращается к неактивной форме - αβγ-ГДФ. При последующей активации рецептора и замене молекулы ГДФ на ГТФ цикл повторяется снова. Таким образом, α-субъединицы G-белков совершают челночное движение, перенося стимулирующий или ингибирующий сигнал от рецептора, который активирован первичным посредником (например, гормоном), на фермент, катализирующий образование вторичного посредника.
Некоторые формы протеинкиназ могут фосфорилировать α-субъединицы G-белков. Фосфорилированная α- субъединица не комплементарна специфическому белку мембраны, например аденилатциклазе или фосфолипазе С, поэтому не может участвовать в передаче сигнала.
Гормоны, взаимодействие которых с рецептором клетки-мишени приводит к образованию цАМФ, действуют через трёхкомпонентную систему, которая включает белок-рецептор, G-белок и фермент аденилатциклазу. Образующийся под действием аденилатциклазы цАМФ активирует протеинкиназу А, фосфорилируюшую ферменты и другие белки (см. раздел 5). Известно более 200 различных G-белков, в структуре которых
обнаружены 3 субъединицы α, β и γ (см. раздел 5). В отсутствие гормона α-субъединица G-белка связана с ГДФ. Образование комплекса гормонрецептора приводит к конформационным изменениям α-субъединицы, замене ГДФ на ГТФ и отщеплению димера βγ от α-ГТФ. В случае рецепторов, сопряжённых с Gs-белком, субъединица αs-ГТФ активирует аденилатциклазу (рис. 11-3).
В случае рецепторов, сопряжённых с Giбелком, субъединица αi-ГТФ ингибирует аденилатциклазу. В таблице 11-4 приведены примеры гормонов, взаимодействие которых с соответствующим рецептором активирует или ингибирует аденилатциклазу.
Другая система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Цитоплазматический домен такого типа рецепторов обладает активностью гуанилатциклазы, которая катализирует реакцию образования цГМФ из ГТФ> (подобно аденилатциклазе). Молекулы цГМФ могут активировать ионные каналы либо активировать цГМФ-за-висимую протеинкиназу G, участвующую в фосфорилировании других белков в клетке. Например, фосфодиэстерааа, которая гидролизует цАМФ до АМФ, активируется в результате фосфорилирования цГМФ-зависимой протеинкиназой.
1. Протеинкиназы А
Протеинкиназы А (цАМФ-стимулируемые) участвуют в аденилатциклазной системе передачи сигнала. Протеинкиназа А состоит из 4 субъединиц R2C2 - двух регуляторных субъединиц (R2) и двух каталитических (С2) (см. рис. 5-41). Комплекс R2C2 нe обладает ферментативной активностью.
Комплекс R2C2 разными способами прикрепляется к мембране. Некоторые формы Протеинкиназы А "заякориваются" с помощью алифатического остатка миристиновой кислоты каталитических субъединиц. Во многих тканях протеинкиназа А связана с "заякоренным" белком AKAPS (от англ. cAMP-dependent protein kinase anchoring proteins). AKAPs имеет центр связывания для регуляторных субъединиц Протеинкиназы А. С помощью белка AKAPS протеинкиназа А связывается с мембраной в области локализации ферментов, катализирующих образование цАМФ (аденилатциклаза) или его гидролиз (фосфодиэс-тераза), а также белков, в регуляции активности которых фермент принимает участие, например потенциалзависимые Са2+- каналы.
Регуляторные субъединицы Протеинкиназы А имеют специфические центры для связывания цАМФ. Присоединение цАМФ к регуляторным субъединицам приводит к изменению конформации последних и снижению сродства к каталитическим субъединицам С, происходит диссоциация по схеме:
цАМФ4 + R2C2 → цАМФ4R2+ С + С
Субъединицы С представляют собой активную форму Протеинкиназы А, которая катализирует реакции фосфорилирования белков по серину и треонину. Каталитические субъединицы С у разных типов протеинкиназ А не идентичны, они различаются прежде всего специфичностью в отношении белков- субстратов.
3.Протеинкиназы G
В отличие от Протеинкиназы А, протеинкиназа G присутствует не во всех тканях, её обнаруживают в лёгких, мозжечке, гладких мышцах и тромбоцитах. Изоформы Протеинкиназы G могут быть связаны с мембраной или находиться в цитоплазме. Растворимая протеинкиназа G состоит из двух идентичных субъединиц, каждая из которых имеет два центра для связывания цГМФ. Присоединение цГМФ к регуляторным центрам вызывает конформационные изменения субъединиц и повышает каталитическую активность фермента (рис. 5-39). Протеинкиназа G, подобно протеинкиназе А и С, специфична в отношении определённых белковых субстратов, которые она фосфорилирует по остаткам серина и треонина.
|
 Скачать 1.15 Mb.
Скачать 1.15 Mb.