Шпоры культура клеток органов и тканей. ККОИТ. 1. История развития метода культивирования клеток и тканей растений
 Скачать 1.38 Mb. Скачать 1.38 Mb.
|

|
7. Среда Андерсона Компоненты/среда Андерсона NH4 NO3 400 Ca(NO3)2 x 4H2 O - MgSO4x 7H2 O 370 NaH2 PO4x H2 O 380 CaCl2x 2H2 O 440 KH2 PO4 - К2 SO 4 - (NH4)2SO4 - (мг/л) Среда используется при введении экспланта в культуру in vitro. Из-за специфичности морфо- и органогенетической реакции сортов растений на активность ростовых веществ, витаминов, макро- и микроэлементов и их композици. Сорта культурных растений нуждаются в индивидуальном подборе компонентов питательных сред и их соотношений для наиболее эффективного управления ростом и развитием эксплантов in vitro. Цитокинины стимулируют у листьев рост в фазе растяжения, а также клеточные деления. Они осуществляют общую стимуляцию обмена веществ, что проявляется в усилении биосинтеза нуклеиновых кислот и белков и значительно замедляют старение клеток, способны вызывать вторичное зеленение пожелтевших листьев. В ряде случаев цитокинины стимулируют образование фенольных веществ и пигментов. Усиление биосинтетических процессов под влиянием цитокининов способствует притоку аминокислот, фосфатов и т.д. к местам, где они локализуются (растущие плоды, семена, клубни), проявляется так называемая аттрагирующая активность цитокининов. Кроме того, цитокинины вызывают заложение и рост стеблевых почек у недифференцированных каллусов. Гиббереллины усиливают рост стеблей за счет вытягивания, но не увеличения числа междоузлий, вызывают прорастание семян и образование партенокарпических плодов, индуцируют стрелкование розеточных форм растений, нарушают период покоя. Стимулируя рост стебля, гиббереллин одновременно способен подавлять рост боковых побегов и корней, уменьшать размеры листьев. Ауксины активируют рост отрезков колеоптилей, стеблей, листьев, корней, вызывают трофические изгибы, стимулируют образование корней у черенков растений. При культивировании микрорастений in vitro наиболее важным у цитокининов является свойство вызывать заложение и рост дополнительных розеток у недифференцированных каллусов, у ауксинов – способность стимулировать образование корней у микрочеренков растений, а у гиббереллинов – усиливать рост стеблей за счет вытягивания. 8-9. Среда Мурасиге и Скуга Наиболее часто используемая среда Мурасиге и Скуга. Минеральная основа этой среды была подобрана авторами для каллусной ткани табака сорта ≪Висконсин≫, возникшей на экспланте – паренхиме стебля в средней его части. Эта среда содержит хорошо сбалансированный состав питательных веществ и отличается от других соотношением аммонийного и нитратного азота. Она подвергается разным модификациям, что реже относится к макро- и микроэлементам минеральной основы и чаще касается набора витаминов, гормональных и других регуляторных факторов. Среда Мурасиге и Скуга дает хорошие результаты при каллусообразовании у большинства растений, хорошо поддерживает неорганизованный каллусный рост и вызывает индукцию морфогенеза у большинства двудольных. 9. Среда Гамборга используется при культивировании клеток и тканей бобовых растений и злаков. 10. Среда Уайта Среда Уайта обеспечивает укоренение побегов и нормальный рост стебля после регенерации. Компоненты Концентрация компонентов в среде (%)  Концентрация компонентов в среде (мг/л)  11. Регуляторы роста и развития растений На рост растений влияют следующие факторы внешней среды: 1. Свет. Органы растений, растущие без света называют этиолированными. При недостатке света у растений происходят глубокие анатомические и физиологические изменения: плохо развиваются механические ткани, устьица, не развита столбчатая паренхима, отсутствуют хлоропласты, выражены длинные междоузлия. При освещении таких органов происходит остановка удлинения междоузлий, формирование механических тканей, синтез хлорофилла и др. Большое влияние на рост оказывает спектральный состав света: а) коротковолновая часть света стимулирует процессы деления клеток, но задерживает фазу растяжения клеток; б) длинноволновая часть подавляет деление клеток, процессы развития и формирования листьев, стебли вытягиваются, отличаются рыхлостью, полное отсутствие механических тканей, усиливается линейный рост клеток. Фотопериодизм –общее свойство всех живых организмов реагировать на суточный ритм освещения, то есть на соотношение светлого и темного периода суток. Фотопериодизм выражается в изменение процессов роста и развития. По особенностям фотопериодизма зеленые растения подразделяются на: 1) растения длинного дня – которым для нормального происхождения всех стадий развития требуется длинный световой день – не менее 12 часов (лен, пшеница и др.); 2) растения короткого дня – полностью развиваются только при коротком световом дне (просо, кукуруза, виноград, хлопчатник); 3) нейтральные растения – развитие которых не зависит от продолжительности светового и темнового периодов (подсолнечник, гречиха, горох). Фотопериодизм – важная адаптация, обеспечивающая заблаговременную подготовку к неблагоприятным зимним условиям, к периоду покоя и зависит от спектрального состава света. 2. Температура. Для растений разных климатических зон характерны свои температурные минимумы, оптимумы и максимумы. Различают три кардинальные точки влияния температур на растения: ► минимальная – точка ниже которой ростовые процессы прекращаются; ► максимальная – точка, выше которой ростовые процессы прекращаются; ► оптимальная – наиболее интенсивно идут процессы роста. Оптимальная температура наиболее благоприятна для роста. У растений умеренной климатической зоны повышение температуры до 30-35 °С вызывает ускорение роста, выше – тормозит. На различных стадиях развития потребность в тепле различна. Устойчивость к низким температурам – важное приспособительное свойство. 3. Газовый состав воздуха. Концентрация кислорода большей частью не влияет на скорость роста, даже при затоплении рост некоторое время продолжается за счет использования кислорода воздухоносных тканей и нитратов. Увеличение концентрации углекислого газа до 0,3% вызывает усиление роста за счет ускорения процессов фотосинтеза, свыше – замедляет и прекращает рост. 4. Водный режим. Только в насыщенных водой клетках нормально протекают все процессы. Недостаток воды подавляет рост, для таких органов характерна мелкоклеточность, длительный избыток воды вызывает остановку роста. 5. Минеральное питание. Азот – ускоряет рост, но задерживает дифференцировку тканей, закладку цветков. Калий, бор существенного влияния на рост не оказывает, но ускоряют цветение. 12. Фитогормоны как природные регуляторы роста и развития растений Фитогормоны (ростовые вещества) – низкомолекулярные органические вещества, образующиеся в различных тканях и органах и, действующие в очень низких концентрациях (10-13-10-5 моль/л). Впервые о возможности существования у растений веществ, сходных с гормонами животных предположил еще Ч. Дарвин в своей книге «Способность к движению у растений» в 1880 г. Он исследовал тропизмы растений и установил, что наиболее чувствительна к воздействию внешних факторов верхушка органа. Это сенсорные органы. Медленные двигательные реакции осуществляются в другом участке – моторной зоне. В связи с этим он предположил, что раздражение, воспринятое сенсорной зоной, передается в моторную с помощью какого-то химического фактора. Значение ростовых веществ или регуляторов роста и развития растений в том, что они позволяют воздействовать на интенсивность и направленность физиологических процессов в растениях, повышать их урожайность, улучшать качество и др. В 1909-1910 гг. Г. Фиттинг обнаружил присутствие гормоноподобных веществ в поллиниях орхидей и назвал эти вещества гормонами. К 60-м годам XX в. были открыты и изучены все группы гормонов, известные на сегодняшний день, которые принимают участие во всех жизненных процессах растений. Гормональная система растений в отличие от животных менее специализирована, и для включения и выключения каких-либо физиологических и морфогенетических программ используются одни и те же гормоны, но в разных концентрациях (Рис.). Фитогормон, также как и гормоны животных, синтезируются в одних органах, а оказывают физиологический эффект на другие органы (рис.) 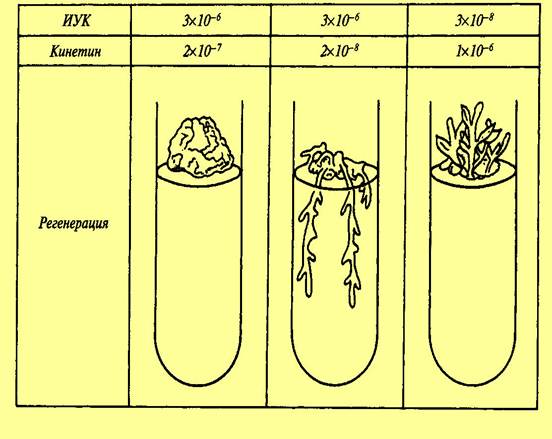 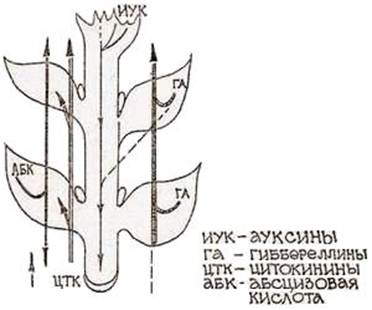 Рис. Зависимость регенерации культуры Рис. Синтез и транспорт фитогормонов ткани сердцевины табака от соотношения фитогормонов. Фитогормоны по механизму действия и химической природе подразделяют на: ► гормоны – стимуляторы (ауксины, цитокинины, гиббереллины) ► гормоны – ингибиторы, тормозящие физиологические процессы (абсцизины и этилен). 13. Ауксины: метаболизм и транспорт, физиологическое действие Ауксины - (от греч. auxano -- расту), группа фитогормонов. Активируют метаболизм, необходимы для роста и развития растений, дифференциации органов, ориентации по отношению к свету и силе тяжести. По химической природе -- производные индола. Исследование эффектов ауксинов начато Ч. Дарвином и его сыном в 1880, продолжено П. Бойсен-Йенсеном (1913), А. Паалем (1919), Н. Г. Холодным и Ф. Вентом (1924-28) и др. Основным ауксином является индолил-3-уксусная кислота (ИУК). Обнаружены и другие природные вещества с ауксиновой активностью, такие как индолил-3-ацетонитрил, 4-хлор-3-индолилуксусная и фенилуксусная кислоты, однако их ауксиновая активность существенно слабее. В растениях часто обнаруживают конъюгаты ИУК с аминокислотами, сахарами и спиртами, представляющие, как полагают, запасные формы ауксинов. Искусственно синтезирован ряд соединений с высокой ауксиновой активностью (2,4-дихлорфеноксиуксусная-, 2,4,5-трихлорфеноксиуксусная-, индолил-3-масляная-, 1-нафтилуксусная кислоты и другие), часто применяемых для научных и практических целей. ИУК синтезируется из триптофана в верхушках побега и перемещается сверху вниз по паренхимным клеткам со скоростью 10-15 мм/час благодаря особому механизму полярного транспорта. Возможно также более быстрое передвижение ауксинов по транспортным каналам растения. Ауксины обладают многообразным физиологическим действием и жизненно важны для роста и развития растений. Ауксины необходимы для деления и растяжения клеток, для формирования проводящих пучков и корней, способствуют разрастанию околоплодника. Ауксины обусловливают явление апикального доминирования, т. е. тормозящее действие апикальной почки на рост пазушных почек. Ауксины играют первостепенную роль в ростовых движениях: фото- и геотропизме и настиях. Ауксины усиливают аттрагирующее действие органов и тканей (т. е. их способность притягивать питательные вещества) и во многих случаях задерживают их старение. При реализации многих физиологических программ ауксины взаимодействуют с цитокининами и другими фитогормонами. При высоких концентрациях ауксины повышают образование своего антагониста -- фитогормона этилена. Первичное действие ауксинов направлено на изменение активности (активацию или репрессию) определенного набора компетентных генов, характерного для данной ткани. ИУК также активирует АТФазу плазмалеммы, вызывая выкачивание протонов из клетки и закисление клеточной стенки. Это приводит к размягчению матрикса стенки, что делает возможным рост клеток растяжением. 14. Цитокинины: метаболизм и транспорт, физиологическое действие Цитокинины - группа фитогормонов, производные азотистого основания пурина, необходимые для деления клеток, роста и дифференцировки растений. Природные цитокинины -- зеатин (6-(4-окси-3-метил-2-бутенил)аминопурин), изопентениладенин, дигидрозеатин, о-оксибензиладенин -- часто выявляются в виде рибозидов и риботидов. Рибозиды считаются основной транспортной формой цитокининов. Цитокинины образуются главным образом в кончиках корней и перемещаются в верхние части растения по ксилеме; заметные количества цитокининов обнаруживаются и во флоэме. Цитокинины типа изопентениладенина и зеатина найдены в составе некоторых тРНК, причем не только у растений. Цитокинины синтезируются многими фитопатогенными и симбиотическими микроорганизмами, что помогает последним воздействовать на клетки растения-хозяина. Цитокинины обладают многообразным физиологическим действием и жизненно важны для роста и развития растений. Совместно с ауксинами они активируют деление клеток, стимулируют развитие боковых побегов (снятие апикального доминирования), в культуре клеток способствуют клеточной дифференцировке и формированию побегов. Цитокинины усиливают способность клеток притягивать питательные вещества (аттрагирующий эффект) и задерживают старение листьев многих растений. Цитокинины активируют формирование хлоропластов и усиливают газообмен растений за счет открывания устьиц. Для многих растений, цитокинины способствуют прорастанию семян и повышают их всхожесть. Цитокинины увеличивают размеры клеток листа и тем самым усиливают рост молодых листьев. Цитокинины обладают и определенным защитным действием на растения против неблагоприятных внешних условий. Действие цитокининов связано с изменением активности (активацией или репрессией) определенного набора компетентных генов, характерного для данной ткани. В ряде случаев цитокинины активируют аппарат синтеза белка независимо от воздействия на геном клетки. На практике используют цитокинины как аденинового ряда (6-бензинаденин, кинетин), так и производные фенилмочевины с высокой цитокининовой активностью (дропп, 4-PU). Цитокинины и их синтетические аналоги применяют для размножения клеток и растений в культуре ткани и для получения трансгенных растений (совместно с ауксинами). Цитокинины используют с целью усиления кущения растений, изменения формы и увеличения размеров ряда плодов, повышения доли женских цветков (у огурца), всхожести семян, устойчивости растений к абиотическим стрессам и болезням. Дропп применяют для предуборочной дефолиации хлопчатника. 15. Фитогормональные стероиды: метаболизм и транспорт, физиологическое действие У растений в-во стероидной природы, обладающее ростостимулирующим эффектом, в чистом виде впервые было получено из липидной фракции пыльцы рапса. Этот первый стероидный фитогормон получил название брассинолида. В наст. время в растит. орг-мах выявлено более 60 схожих с брассинолидом в-в, называемых брассиностероидами (БС), к-ые синтезир-ся гл. обр. из кампестерола. Больше всего БС сод-т генеративные органы растений. Одной из особенностей БС, отличающей их от других фитогормонов, является спос-ть стимулир-ть ростовые процессы в очень низких конц-иях (10-6-10-12 М). Однако следует отметить, что обработка БС не всегда вызывает полож. эффект в биотестах, характерных для ИУК, гиббереллинов и цитокининов. Наиболее специфичный для БС эффект заключ-ся в активации роста 2-го междоузлия фасоли / гороха. Ни ауксины, ни цитокинины не вызывают такого эффекта, а гиббереллины активируют удлинение междоузлий фасоли только в очень высокой конц-ии. Еще более чувствительным биотестом на БС явл-ся рост эпикотилей маша. На этом объекте стимулир-ий эффект БС проявляется в конц-ии 10-10 М и ниже. Стимулир-ее действие БС на ростовые процессы связано с активацией процессов деления и растяжения клеток. Поэтому в отличие от гиббереллинов БС вызывают не только удлинение 2-го междоузлия ряда бобовых растений, но и его утолщение за счет активации клеточных делений. БС участвуют в регуляции ростовых процессов во взаимод-ии с др. фи-тогормонами. Выявлено, что БС и гиббереллины оказывают аддитивное действие на рост гипокотилей огурца, к-ые особенно чувствительны к брассинолиду. Обнаружен сильный синергический эффект ИУК и брассинолида, к-ый проявлялся только в том случае, когда растения предварительно обрабатывались БС, а затем ИУК. Если же обработка растений шла в иной послед-ти, синергизма не наблюдалось. Ингибитор тр-та ауксина 2,3,5-трииодбензойная кислота устраняла синергический эффект. Экзогенная обработка БС в низких конц-иях оказывает влияние не только на процессы роста и разв-ия растений, она может повышать их уст-ть к таким стрессовым возд-иям, как резкие колебания температуры, засуха, засоление, аноксия и возд-ие патогенов. Поэтому БС рассматриваются как одна из наиболее перспективных групп эндогенных фитогормонов для растениеводства, поскольку их можно исп-ть в ничтожных кол-вах не только для регулир-ия процессов роста и разв-ия, но и для защиты растений. При исп-ии в с/х наиболее эффективны комбинации БС с др. фитогормонами — ГК, АБК, этиленом, цитокининами. БС играют важную роль в процессах фотоморфогенеза. Есть предположение, что они влияют на акт-ть ряда генов, экспрессия к-ых индуцируется при попадании этиолированных проростков на свет. Очень интересные в этом плане рез-ты получили в 2000 г. Нагата с коллегами при исп-ии ингибитора синтеза БС — брассиназола. После обработки брассиназолом этиолированные проростки арабидопсиса приобретали хар-ки растений, растущих на свету: широкие семядоли, короткий гипокотиль, истинные листья, в семядолях начиналось формирование пластид и повышался уровень РУБИСКО. Поэтому полагают, что БС выполняют ф-ию репрессора светозависимых генов морфогенеза. Выявлено 2 типа брассиностероидных мутантов. К 1-му типу относят мутан-ты, у к-ых нарушены разл. этапы синтеза БС. К ним относятся, например, 2 фотоморфогенетических мутанта арабидопсиса: det2 (de-etiolated), и сpd (constitutive photomorphogenesis dwarfism). У этих мутантов в условиях темноты не проявляется явление этиоляции — растения имеют развернутые семядоли, не вытянутый, а короткий побег и дифференцированные хлоропласты. Обработка таких мутантов экзогеными БС восстанавливает норм. фенотип растения. Ген. анализ мутантов с нарушением этиоляции показал, что Мц затрагивают гены DET2 и CPD, кодир-ие ферменты синтеза БС. Мутация det2 затрагивает ген DET2, к-ый кодирует белок, гомологичный одному из ключевых ферментов метаболизма стероидов — кампестерол-5-редуктазе, катализир-ей восстановление кампестерола в кампестанол. Мутация cpd блокирует гидроксилирование С21 в молекуле катастерона и превращение его в тиастерон. Поэтому обработка мутантов det2 и cpd экзогенными брассиностероидами восстанавливает норм. фенотип растений. Изучение плейотропных эффектов мутаций det2 и cpd у арабидопсиса показало, что БС играют важную роль в регуляции таких свето- и гормон-регулируемых процессов, как экспрессия светорегулируемых генов, рост клеток растяжением, старение листьев, индукция цветения. 2-й тип брассиностероидных мутантов связан с нарушением процессов рецепции и (или) передачи гормон. сигнала, поэтому обработка экзогенным гормоном не оказывает на них влияния. Ген. анализ мутантов, нечувствит-ых к обработке БС, показал, что Мц затрагивают один и тот же ген — BRI1, к-ый кодирует рецепторную серин/треониновую протеинкиназу с лейцин-богатыми повторами — LRR-RLK — от англ. leucine rich repeat receptor-like kinase. Установлено, что рецептор брассиностероидов BRI1 состоит из 1196 остатков АК, локализован в плазмат. мембране и имеет типичную стр-ру LRR-RL-киназы, широко распространенной у растительных и животных орг-мов. |