Глик Молекулярная биотехнология. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение. Пер с англ. М. Мир, 2002. 589 с
 Скачать 9.74 Mb. Скачать 9.74 Mb.
|

ГЛАВА 6. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
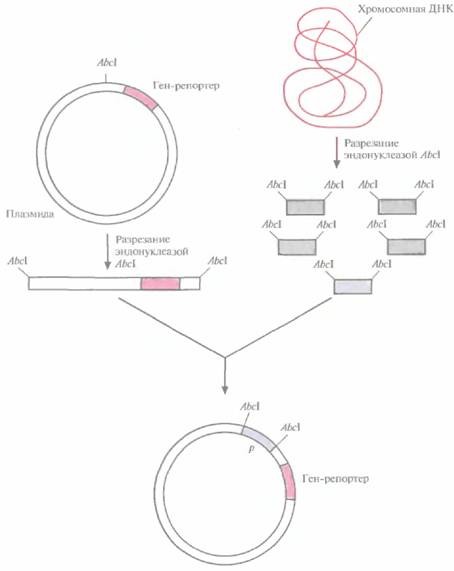 | Рис. 6.1. Идентификация сильных регулируемых промоторов. В плазмиду встраивают ген-репортер без промотора. Хромосомную ДНК. разрезают рестрицируюшей эндонуклеазой AbcIи встраивают фрагменты в плазмиду. Если ген-репортер эффективно экспрессируется, значит, клонированный фрагмент содержит функциональный промотор. |
Оптимизация экспрессии генов, клонированных в прокариотических системах 107
в культуре преобладающими. Нестабильность плазмид — это основная проблема, мешающая получению продукта гена, локализованного в плазмиде, в промышленных масштабах. Для ее решения нужно научиться контролировать экспрессию таким образом, чтобы клонированный ген экспрессировался только в определенной фазе клеточного цикла и только в течение определенного времени, а для этого нужно использовать сильные регулируемые промоторы. Плазмиды, сконструированные для этих целей, называются экспрессирующими векторами.
Регулируемые промоторы
Наиболее широко используются следующие сильные регулируемые промоторы: промоторы lac- и trp-оперонов E. соli; специально сконструированный tac-промотор, включающий — 10-область lас-промотора и — 35-область trp-промотора (участки, находящиеся на расстоянии 10 и 35 п. н. до сайта инициации транскрипции); левый, или pL, промотор бактериофага λ; промотор гена 10 бактериофага Т7. С каждым из них связываются соответствующие репрессоры, которые опосредуют включение и выключение транскрипции специфических генов. Кроме того, каждый из этих промоторов узнается холо-ферментом РНК-полимеразой E. coli, в который входит основной сигма-фактор, присутствующий в клетке в значительно больших количествах, чем другие, минорные сигма-факторы. Благодаря этому транскрипция не останавливается по причине недостатка свободных сигмафакторов.
В отсутствие лактозы в среде lас-промотор E. coli находится в репрессированном состоянии, т. е. он выключен белком-репрессором, блокирующим транскрипцию lac-оперона. Индукция, или включение lac-оперона происходит при добавлении в среду лактозы или изопропил-β-D-тиогалактопиранозида (ИПТГ). Оба этих соединения предотвращают связывание репрессора с lac-оператором, и транскрипция возобновляется.
Транскрипция, контролируемая lac-промотором, регулируется также с помощью белка -активатора катаболизма (САР) (рис. 6.2). При связывании САР с промотором повышается сродство последнего к РНК-полимеразе и усиливается транскрипция примыкающих к нему генов. В свою очередь сродство САР к промотору повышается при его связывании с цикличе-
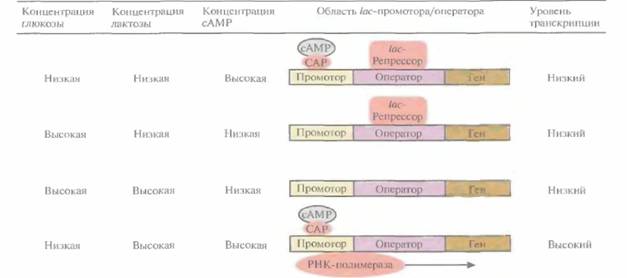 |
| Рис. 6,2. Влияние глюкозы, лактозы и сАМР на транскрипцию, регулируемую lac-промотором E. coli. Стрелка указывает направление транскрипции. (По данным работы Abeles et al., 1992, Biochemistry, p. 383, Jones and Bariett Publishers, Boston, Mass.) |
108 ГЛАВА 6
ским AMP (сАМР), уровень которого повышается при снижении концентрации глюкозы в среде. Таким образом, если репрессор не связан с оператором, то в присутствии индуктора при повышении внутриклеточной концентрации сАМP может произойти усиление транскрипции генов, регулируемых laс-промотором.
На самом деле в плазмидных экспрессируюших векторах используется один из вариантов lac-промотора — lacUV5 сизмененной -10-последовательностью, более сильный, чем laс-промотор дикого типа. Транскрипция с промотора tac также подавляется lac-репрессором и возобновляется при добавлении в среду лактозы или ИПТГ.
Промотор trp выключается под действием комплекса триптофан—trp-репрессор, который связывается с trp-оператором и предотвращает транскрипцию trp-оперона. Активация (включение) trp-промотора происходит либо при удалении из среды триптофана, либо при добавлении 3-индолилакриловой кислоты.
Работа промотора рL- регулируется репрессорным белком cl бактериофага λ. На самом деле для регуляции транскрипиии с pL-промотора обычно используется термочувствительная му-тантная форма репрессора cI — белок сI857. Клетки, синтезирующие этот репрессор, сначала выращивают при температуре 28—30 °С; в этих условиях репрессор блокирует транскрипцию с pL-промотора. Когда культура достигает нужной фазы (как правило, середины log-фазы), температуру повышают до 42 °С, при которой сI857-репрессор инактивируется и начинается транскрипция.
Для транскрипции с промотора бактериофага Т7 нужна соответствующая РНК-полимераза. Чтобы можно было использовать этот промотор, ген РНК-полимеразы фага Т7 встраивают в хромосому E. coli в составе профага λ, поместив его подконтроль lac-промотора. Затем клетки трансформируют плазмидой, содержащей генмишень под контролем Т7-промотора, и добавляют в среду ИПТГ. В этих условиях происходит индукция гена РНК-полимеразы Т7, синтезируется РНК-полимераза и происходят транскрипция и трансляция клонированного гена. Часто между временем индукции гена РНК-полимеразы Т7 и началом транскрипции гена-мишени проходит более часа. Для транскрипции с сильного Т7-промотора создана целая серия плазмид, получивших название рЕТ-векторов.
Эффективность инактивации белка-репрессора и соответственно активации транскрипции зависит от соотношения между числом молекул репрессора и числом копий промотора. Если концентрация репрессора слишком велика, то транскрипция не инициируется, и наоборот, если молекул репрессора очень мало (даже при том, что их больше, чем копий промотора), то транскрипция может идти и в отсутствие индукции. Про такие промоторы говорят, что они «текут». Чтобы осуществлять строгий контроль таких регулируемых систем, разработаны разные стратегии. Например, ген репрессора и соответствующий промотор помещают в две разные плазмиды, присутствующие в клетке в разном числе копий; это позволяет поддерживать нужное соотношение между числом молекул репрессора и числом копий промотора. Обычно ген репрессора находится в малокопийной плаз-миде, число ее копий в клетке не превышает 8, а промотор -- в мультикопийной плазмиде с 30—100 копиями на клетку. Ген репрессора может быть локализован и в хромосомной ДНК, находясь в ней в единственном числе, что позволяет поддерживать низкую концентрацию репрессора. В системах, использующих laс-промо-тор, можно получить lac-репрессор в значительно большем количестве, если заменить lacI-ген его мутантной формой laсIq, что приводит к уменьшению «протекания» промотора, т. е. к снижению уровня транскрипции клонированного гена без индуктора.
Получение больших количеств белковых продуктов
Для получения больших количеств чужеродных белков с помощью рекомбинантных штаммов E. coli была сконструирована плазмида pPLc2833. Она содержит сильный промотор, селективный маркерный ген и короткий участок с несколькими уникальными сайтами для рестрицирующих ферментов (полилинкер), следующий непосредственно за промотором. Эффективность этого экспрессируюшего вектора в осуществлении синтеза чужеродных белков в Е. соli можно еще
Оптимизация экспрессии генов, клонированных в прокариотических системах 109
| Таблица 6. I. Зависимость числа копий трех плазмидных экспрессирующих векторов от температуры1) |
 |
больше повысить, заменив сайт инициации репликации плазмиды pPLc2833 аналогичным сайтом плазмиды рКМ402. Это приводит к увеличению числа копий модифицированной плазмиды в 5-10 раз при температуре 42 °С (табл. 6.1), Полученная таким образом плазмида рСРЗ содержит pL-промотор и ген ß-лактамазы (ген устойчивости к ампициллину) из pPLc2833, a сайт инициации репликации - из pKN402 (рис. 6.3). Несущие ее клетки сначала выращивают при температуре 28 °С, а затем — при 42 °С. При пониженной температуре ген cI-penpeccopa, интегрированный в ДНК E. соli, экспрессируется, pL-npoмотор не функционирует и образуется обычное число копий плазмиды (табл. 6.1). При повышении температуры cl-penpecop инактивируется, pL-промотор переходит в активное состояние, и число копий плазмиды увеличивается. Все это и делает плазмиду рСР3 эффективным экспрессирующим вектором. Когда ген ДНК-лигазы Т4 был встроен в полилинкер рСР3, то выход его продукта составил примерно 20% от общего количества белка, синтезируемого Е. соli при 42 °С. При этом на долю собственных, наиболее активно синтезируемых белков E, coli, например фактора элонгации EF-Тu, приходится примерно 2%.
Крупномасштабные системы
При культивировании в небольших объемах (от l до 5 л) индукцию экспрессии осуществляют либо изменением температуры, либо добавлени-
| Рис. 6.3. Создание плазмиды рСР3. Из плазмиды pKN402 с помощью рестрицирующей эндонуклеазы HaeIIвырезают фрагмент с, содержащий температурочувствительный сайт инициации репликации (ori), и сшивают его с НаeII-фрагментами 1 и 3 плазмиды pPLc2833. Фрагмент 1 содержит pL--промотор и полилинкер (ПЛ), а фрагмент 3 - селективный маркерный ген устойчивости к ампициллину (Ampr). | 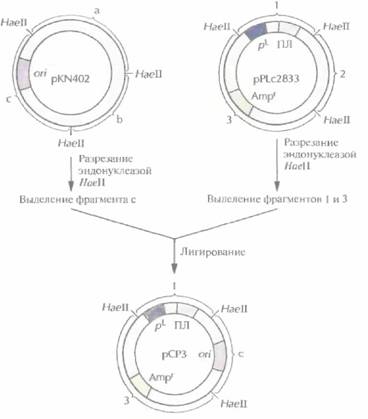 |
110 ГЛАВА 6
ем химического индуктора. Однако в опытных установках (20—100 л) и в промышленных биореакторах (>200 л) температуру нельзя изменить мгновенно, для этого требуется время от 30 до 60 мин; кроме того, на подъем температуры нужна энергия. И время, и энергия стоят дорого. Столь же дорого обходится и применение химического индуктора, например ИПТГ. Все это может сделать процесс неэкономичным. Для преодоления некоторых проблем, связанных с использованием pL-промотора для крупномасштабного производства белковых продуктов, была разработана двухплазмидная система. Ген репрессора cI поместили под контроль trр-про-мотора и включили в малокопийную плазмиду (рис. 6.4), что обеспечило невысокий уровень синтеза репрессора. Вторая плазмида содержала клонированный ген, находящийся под контролем pL-промотора. Как видно из рис. 6.4, А, в отсутствие триптофана включается trр-промотор и синтезируется репрессор cI, выключающий pL-промотор. И наоборот, как видно из рис. 6.4, Б, при наличии триптофана trp-промотор выключается, репрессор не синтезируется, а pL-промотор активно работает.
Культуры с такими двухплазмидными системами можно выращивать на недорогих средах на основе гидролизатов мелассы или казеина, содержащих незначительное количество свободного триптофана, и индуцировать экспрессию клонированного гена добавлением в среду трип-тона. Последний содержит свободный триптофан в количестве, достаточном для эффективной индукции транскрипции. Пробные испытания этой системы показали, что на долю продуктов клонированных генов ß-лактамазы и цитратсин-
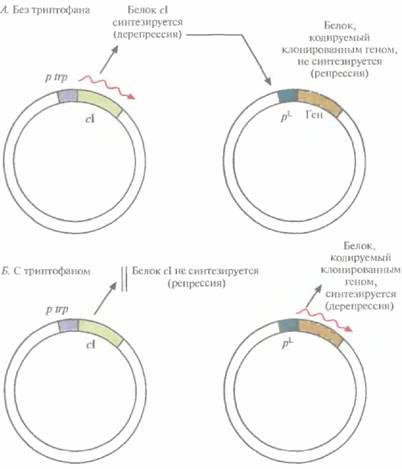 | Рис. 6.4. Двухплазмидная система, позволяющая контролировать работу pL-промотора фага λ путем регуляции синтеза cl-peпрессора с помощью триптофана. Ген репрессора cI вместе с триптофановым промотором (р trp) находятся в одной плазмиде, а pL-промотор и клонированный ген — в другой. Стрелками указано направление транскрипции. А. В отсутствие триптофана в среде ген cI транскрибируется и транслируется, репрессор cI связывается с pL-промотором и блокирует транскрипцию клонированного гена. Б. В присутствии триптофана ген cI репрессируется, его продукт не синтезируется, поэтому клонированный ген транскрибируется и транслируется. |
Оптимизация экспрессии генов, клонированных в прокариотических системах 111
тазы после индукции транскрипции добавлением триптона приходится соответственно 21 и 24% от общего количества синтезируемого белка. Таким образом, двухплазмидные системы позволяют получать белковые продукты с помощью реком-бинантных микроорганизмов в промышленных масштабах и относительно недорого.
Использование для экспрессии других микроорганизмов
Е. coll — не единственный микроорганизм, который используется для синтеза чужеродных белков. К сожалению, генетические и молекулярно-биологические свойства большинства других микроорганизмов изучены не так хорошо. Кроме того, нет ни одного вектора или даже промо-торно-репрессор ной системы, которая обеспечивала бы оптимальный уровень экспрессии в клетках всех или хотя бы только грамотрицательных бактерий. К счастью, многие стратегии, разработанные для Е. coli, пригодны и для множества других микроорганизмов, что позволило проверить различные промоторы на их способность обеспечивать транскрипцию в других гра-мотрицательных бактериях. Так, в одном из исследований был сконструирован набор плазмидных экспрессирующих векторов, содержащих промоторы lас, tac, Nm (гена устойчивости к неомицину) и SI (гена рибосомного белка S1 Rhisobium meliloti), и определен уровень экспрессии гена ß-лактамазы под контролем каждого из них (табл. 6.2). Обнаружилось, что: 1) указанные промоторы проявляют ту или иную активность во всех использованных бактериальных системах; 2) промотор tac наиболее активен в Е. coli и гораздо менее — в других бактериях; 3) Nm — второй по активности промотор в E. coli и самый активный в других бактериях. Промоторные участки у всех грамотрицательных бактерий имеют сходную нуклеотидную последовательность, однако это не означает, что самым эффективным промотором для того или иного организма будет тот, который наиболее эффективен в Е. coli. Тем не менее Е. соli-промоторы могут оказаться вполне приемлемыми для регуляции экспрессии клонированных генов и в других грамотрицательных бактериях.
Попытки создания «универсального» экспрессирующего вектора для грамотрицательных бактерий были весьма многочисленными. В конце концов была выбрана следующая стратегия. Фрагмент ДНК размером 70 п. н., происходящий от одного из концевых инвертированных повторов в транспозоне 5 (Tn5), встроили вместе с соответствующим промотором в полилинкер малокопийной плазмиды pRK290 с широким спектром хозяев и получили плазмиду pAV10 (рис. 6.5). Клонированный сегмент ДНК из Tn5 содержал два независимых, но перекрывающихся промотора, каждый из которых необходим для транскрипции одного из ключевых Tn5-генов. Поскольку Tn5 эффективно экспрессируется в разных бактериях, его промоторы можно использовать для транскрипции различных генов. Чтобы проверить это, в полилинкер сразу за клонированными промоторами Тn5 встроили гены хлорамфеникол-ацетил-трансферазы и ß-галактозидазы. Их эффективная экспрессия происходила в клетках Alcaligenes sp,, E. coli, Enterobactercloacae, Klebsiella pneumoniae, Pseudomonas stuizeti, Pseudomonas fluorescens и Serratia marcescens. Таким образом, есть реаль-
| Таблица 6.2.Активность ß-галактозидазы в грамотрицательных бактериях, несущих плазмидный вектор с геном lacZ E.coli игетерологичным промотором1) | ||||
| Промотор | Активность ß-галактозидазы, ЕД | |||
| | Escherichia coli | Rhizobium meliloti | Rhizobium leguminosarum | Pseudomonas putida |
| Отсутствует | 16 | 110 | 130 | 150 |
| Nm | 1400 | 21800 | 13900 | 16300 |
| lac | 2000 | 9050 | 6250 | 9800 |
| tac | 11300 | 2850 | 1150 | 2950 |
| S1 | 40 | 3300 | 1200 | 3350 |
| 1) По данным Labes et al., 1990, Gene 89: 37—46. | ||||
112 ГЛАВА 6
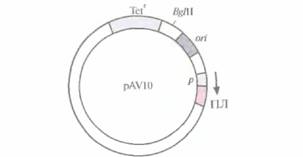 |
| Рис. 6.5. Клонируюший вектор pAV10 (без соблюдения масштаба). Показано положение гена устойчивости к тетрациклину (Tetr), сайта рестрикции для эндонуклеазы BglII, сайта инициации репликации (on), промотора (p) и полилинкера (ПЛ). Встраивание клонированного гена в полилинкер ставит его под контроль промотора Тn5 (р). Стрелка указывает направление транскрипции. |
ная возможность использовать Тn5-промоторы для инициации транскрипции чужеродных генов в клетках различных бактерий.