Конспект лекций по экологии для заочников. Лекция Краткая история и предмет экологии
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Зональное распределение В любое данное время виды внутри сообщества могут быть пространственно распределены в соответствии с различиями окружающей физической среды. Это называется зональным распределением. Хорошим примером служит вертикальная зональность с увеличением высоты в горах. Внешне зональное распределение может иметь сходство с сукцессиями, но необходимо понимать основное различие: при зональном распределении виды сменяются в пространстве, тогда как в сукцессиях они сменяются во времени. Впервые теория сукцессии была детально разработана в 1916 г. Клементсом. Он изучал сообщества в Северной Америке и пришел к выводу, что основным фактором, определяющим состав климаксного сообщества, является климат. По представлениям Клементса, в данных климатических условиях может существовать только одно климаксное сообщество, которое называется климаксом (концепция моноклимакса). Любой перерыв в движении по направлению к климаксу, вызванный местными условиями рельефа, микроклимата, землепользованием и т. д., тоже может привести к формированию устойчивого сообщества, но его нельзя считать климаксным, так как в конце концов действие климатических факторов все-таки приведет к "теоретическому" климаксу. Например, горы и холмы постепенно будут разрушаться, что вызовет изменения в структуре сообществ. Более современной является концепция поликлимакса, согласно которой климакс формируется под влиянием всех физических факторов, причем один или несколько из них могут доминировать (например, дренаж, почва, топография, пожары). Сообщество считается настоящим климаксом, если оно "устойчиво" длительный период времени: любые изменения в нем происходят относительно медленно по сравнению со временем, необходимым для прохождения сукцессии до стадии климакса. Основные особенности сукцессий Предпринималось несколько попыток найти и понять основной принцип, предопределяющий ход сукцессии по направлению к климаксу. Экологическое доминирование Наблюдения, касающиеся экологического доминирования, наводят на мысль, что сукцессия всегда идет в направлении к климаксу, где доминирует один или несколько видов - кодоминантов, например дубовому лесу или к дубово-ясеневому лесу. В таких лесах в связи с затенением и другими причинами ограничен рост видов, которые могли бы существовать рядом с доминантами. Однако в некоторых случаях трудно применить концепцию доминирования, например когда речь идет о сообществах планктона, о животных или о тропических дождевых лесах, где в приблизительно равных количествах можно найти несколько сотен видов деревьев. Продуктивность и биомасса Линдеман в 1942 г. высказал предположение, что сукцессии сопровождаются повышением продуктивности вплоть до климаксного сообщества, в котором превращение энергии происходит наиболее эффективно. Это логичная и привлекательная гипотеза, так как поток энергии, проходящий через экосистему, является одним из важных факторов, лимитирующих численность и биомассу организмов, жизнь которых она способна поддерживать. Данные исследований показывают, что на поздних стадиях сукцессий продуктивность действительно растает, однако при переходе к климаксному сообществу обычно происходит снижение общей продуктивности. Таким образом, продуктивность в старых лесах ниже, чем в молодых, которые в свою очередь могут иметь меньшую продуктивность, чем предшествовавшие им более богатые видами ярусы травянистых растений. Сходное падение продуктивности наблюдается в некоторых водных системах. Как сейчас полагают, во многих сукцессиях максимум продуктивности достигается очень быстро, хотя данных об этом пока еще мало. Причины снижения продуктивности остаются предметом гипотез, хотя, если рассматривать в качестве примера лес, можно ожидать, что продуктивность старых деревьев действительно будет меньше, чем молодых. Для этого есть несколько причин. Одна из них то, что накопление питательных веществ в растущей биомассе леса на корню может вести к уменьшению их круговорота. Однако снижение общей продуктивности могло быть просто результатом уменьшения жизненности особей по мере увеличения их среднего возраста в сообществе. Хотя нет, по-видимому, оснований утверждать, что сукцессии ведут к максимальной эффективности превращения энергии (к максимуму общей продуктивности), уже можно сказать, что они ведут к максимальному накоплению биомассы. Это наиболее очевидно в лесных сообществах, где в ходе сукцессии растения становятся все крупнее и крупнее. Накопленная биомасса в других климаксных сообществах тоже обычно больше, чем в период сукцессии. Изменения в общей продуктивности, дыхании и биомассе в ходе типичной сукцессии показаны на рис. 15.1. 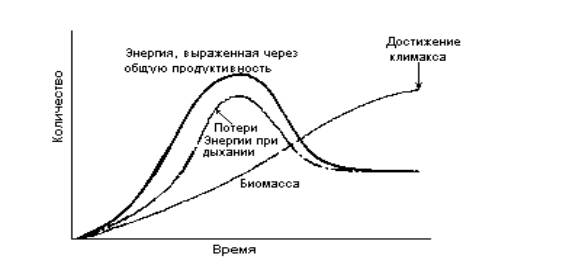 Рис.15.1. Изменения общей продуктивности, дыхания и биомассы в ходе типичной сукцессии. Можно видеть, что в климаксном сообществе эти параметры становятся более или менее постоянными; видно также, что верхний предел накопления биомассы достигается тогда, когда общие потери на дыхание (R) становятся почти равными общей первичной продуктивности (Р), т.е. отношение Р/R приближается к единице. Эти и другие особенности, вероятно, свойственные сукцессиям, приведены в табл. 15.1. Таблица 15.1 Основные изменения в экосистеме во время типичной вторичной сукцессии
По мере прохождения сукцессии все большая доля доступных питательных веществ накапливается в биомассе сообщества и соответственно уменьшается их содержание в абиотическом компоненте экосистемы (в почве или воде). Возрастает также количество образующегося детрита, и главными первичными консументами становятся не травоядные, а детритоядные организмы. Соответствующие изменения происходят и в трофических сетях, и детрит становится основным источником питательных веществ. Взаимодействия между организмами и абиотической средой на ранних стадиях сукцессии наиболее важны взаимодействия между живыми организмами и абиотической средой. В результате деятельности организмов эта среда с течением времени изменяется. Примером может служить образование песчаных дюн вследствие накопления эолового песка вокруг побегов песколюба (Ammophila, или Psamma). Время стабилизации дюн составляет несколько лет. Хорошо изучена сукцессия песчаных дюн вокруг озера Мичиган в Северной Америке, где стабилизация дюн привела к появлению более крупных и не требовательных к питательным веществам растений (тополь, сосны). За длительный период времени образуется почва, в которой накапливается органический углерод, поступающий из подстилки. Увеличение численности азотфиксирующих организмов приводит к росту количества азота в почве. Местами здесь появился дуб, который стал доминирующим видом. После того как дуб окончательно закрепился, почвенные условия, по-видимому, остаются постоянными в течение неопределенного времени: потребность в питательных веществах удовлетворяется за счет поступления их из лесной подстилки. В этом примере почвенные факторы играют определяющую роль в сукцессии и сами изменяются в процессе сукцессионных смен. Может, однако, потребоваться тысяча лет, для того чтобы химический состав почвы стабилизировался и сукцессию можно было считать законченной. Этот пример иллюстрирует важный общий принцип: ранние стадии сукцессии проходят значительно быстрее, чем поздние. Дальнейшее развитие климаксного сообщества происходит только при изменении условий окружающей среды. Хорошим примером изменений климата, приведших к коренной смене сообществ на больших площадях, служит последовательность ледниковых периодов в последние 1,8 млн. лет. Биотическое сообщество может влиять на физическую среду, а тем самым и на сукцессию различными способами. Например, сообщества могут локально изменять климат и формировать микроклимат. Аналогичным образом создаются микроместообитания, такие, как отмершая древесина (в которой может обитать около 200 видов животных), навоз (более 300 видов) и падаль. Взаимодействия между организмами внутри сообщества На более поздних стадиях сукцессий повышается роль биотических взаимодействий в формировании детальной структуры сообщества. Возрастает разнообразие живых организмов, и их взаимодействия неизбежно становятся более сложными и многообразными. Все доступные экологические ниши постепенно заполняются и может возникнуть конкуренция за какую-либо нишу. Шансы на то, что какой-то вид животных закрепится в данной области, зависят от многих причин - от вероятности миграции его в данный район, от наличия пригодной для него пищи, от его способности отыскать подходящую нишу и в случае необходимости защитить ее от конкурентов. Чем больше специализирован вид для использования данной ниши, тем меньше вероятность прямой конкуренции. С представлением об экологических нишах тесно связано понятие разделения ресурсов, т. е. распределения их между различными видами сообщества. Специализация видов в отношении ресурсов уменьшает конкуренцию и увеличивает стабильность структуры сообщества. Существуют различные типы разделения ресурсов: 1) Стабилизация морфологии и поведения в соответствии с родом пищи, например клюв у птиц может быть приспособлен для ловли насекомых, долбления отверстий, раскалывания орехов, разрывания мяса и т. п. 2) Вертикальное разделение, как, например, между обитателями полога и лесной подстилки. 3) Горизонтальное разделение, например, между обитателями различных микроместообитаний. Каждый из этих типов или их комбинация приводят к разделению организмов на группы, которые меньше конкурируют между собой, так как каждая из них занимает свою нишу. Существует, например, разделение птиц на экологические группы, основанные на месте их питания (воздух, листва, ствол, земля). У растений для каждого вида существует набор оптимальных, наиболее благоприятных условий (влажность, высота над уровнем моря, рН, почва и т.д.), что опять-таки приводит к уменьшению прямой конкуренции. Несмотря на то что каждый вид стремится обзавестись собственной нишей, некоторая прямая конкуренция между видами за доступные ресурсы неизбежна. Возрастает роль других типов взаимодействий. Большое значение имеют паразитические, симбиотические и другие мутуалистические отношения. Исследование таких взаимоотношений пересекается с изучением экологии популяций. Лекция 16. Популяционная экология Популяция - это группа особей одного вида (или иные группы, в которых организмы могут обмениваться генетической информацией), занимающая определенное пространство в данное время. Популяция обладает "биологическими особенностями", которые она разделяет со всеми составляющими ее организмами, и "групповыми особенностями", которые служат уникальными характеристиками группы. К "биологическим особенностям" следует отнести наличие жизненного цикла популяции, ее способность к росту, дифференцировке и самоподдержанию. Популяция имеет определенную организацию и структуру, которые можно описать. К "групповым особенностям" относятся такие, как рождаемость, смертность, возрастная структура популяции и генетическая приспособленность; эти признаки относятся только к популяции. Особь рождается и умирает, имеет тот или иной возраст, но применительно к особи нельзя говорить о рождаемости, смертности или возрастной структуре - характеристиках, имеющих смысл только на уровне группы. Следовательно, популяционные признаки можно разделить на две категории: 1) признаки, связанные с количественными соотношениями и структурой: плотность, рождаемость, смертность, распределение организмов по возрастам, биотический потенциал, характер распределения в пределах территории и тип роста. Эти признаки обычно выражаются в виде статистических функций и характеризуют группу как целое; 2) признаки, характеризующие общегенетические особенности популяции, а именно: способностью к адаптации, репродуктивной (дарвиновской) приспособленностью и устойчивостью, т. е. способностью в течение длительного времени производить потомство. Свойства популяционной группы. Плотность популяции. Плотность популяции - это ее величина по отношению к единице пространства. Обычно ее определяют и выражают числом особей или биомассой популяции на единицу площади или объема (например, 200 деревьев на 1 га, 5 млн. диатомовых водорослей на 1 м3 воды, 200 кг рыбы на 1 га поверхности водоема). Подобно некоторым другим популяционным признакам, плотность популяции весьма изменчива. Однако эта изменчивость отнюдь не бесконечна; существуют определенные верхние и нижние пределы для размеров популяции, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени. Так, на значительной площади леса может жить в среднем 100 птиц на 1 га и 20 000 почвенных членистоногих на 1 м2, но никогда не бывает 20000 птиц на 1 м2 и всего 10 членистоногих на 1 га. Верхний предел плотности определяется потоком энергии в экосистеме (продуктивностью), трофическим уровнем, к которому относится организм, а также величиной и интенсивностью его метаболизма. Нижний предел не может быть так хорошо определен, но по крайней мере в стабильных экосистемах действуют гомеостатические механизмы, поддерживающие плотность обычных или доминирующих организмов в довольно ограниченных пределах. Динамика популяций. Скорость роста популяций. Поскольку популяция изменчива, интерес представляет не только ее величина и состав в каждый данный момент, но также и то, как она изменяется. Зная скорость изменения популяции, можно судить о многих ее важных особенностях. Скорость роста популяции - это число организмов, на которое она изменяется за некоторое время; ее получают путем деления величины прироста популяции на протекший промежуток времени. Если через N обозначить. Число организмов, а через t - время, то: DN - изменение числа организмов; DN/Dt - средняя скорость изменения числа организмов во времени (т.е. изменение их числа, деленное на время, это и есть скорость. роста); DN/(NDt) - средняя скорость изменения числа организмов во времени на организм (скорость роста, деленная либо на число организмов. имевшихся вначале, либо на среднее число организмов в данный период времени); часто эту величину называют специфической скоростью роста; ее целесообразно использовать в тех случаях, когда сравниваются популяции разных размеров; если умножить ее на 100 [DN/(NDt)·100], получим скорость роста в процентах. Мгновенные скорости обозначают как dN/dt. dN/dt - скорость изменения числа организмов в зависимости от времени в определенный момент (теоретическая мгновенная скорость в определенное время) - наклон кривой роста популяции в любой ее точке; dN/(Ndt) - скорость изменения числа организмов в зависимости от времени на одну особь в определенный момент. Пример Предположим, что популяция из 50 представителей простейших в некотором объеме воды увеличивается за счет деления. Примем, что через 1 ч она увеличилась до 150 особей. Тогда N (число особей вначале)= 50; DN (изменение численности) = 100; DN/Dt (средняя скорость изменения во времени) = 100 особей в час; DN/(NDt) (средняя скорость изменения во времени на одну особь)= 2 в час на особь (т. е. увеличение за час на 200%). Мгновенную скорость dN/dt, а также скорость dN/(Ndt) нельзя рассчитать непосредственно, лишь на основании подсчетов численности; необходимо также знать характер кривой роста популяции. Рождаемость. Рождаемость - это способность популяции к увеличению. Она характеризует частоту появления новых особей любого организма независимо от того, "рождаются" ли они, "вылупляются", "отпочковываются", "возникают путем деления" и т. п. Термин экологическая, или реализуемая, рождаемость (или просто "рождаемость" без какого-либо определения) обозначает увеличение популяции при фактических или специфических условиях среды. Эта величина изменяется в зависимости от размера и состава популяции и физических условий среды. Максимальная рождаемость (иногда ее называют абсолютной или физиологической) - образование теоретически максимально возможного количества новых особей в идеальных условиях (когда отсутствуют лимитирующие экологические факторы и размножение ограничивается только физиологическими); для данной популяции это величина постоянная. Обычно рождаемость выражают в виде скорости, определяемой путем деления числа вновь образовавшихся особей на время (DNn /Dt, абсолютная рождаемость), или же числом новых особей в единицу времени на единицу популяции ([DNn/(NDt)]), специфическая рождаемость). Рождаемость можно измерить и выразить различными способами. Если через DNn обозначить число особей, вновь образовавшихся в популяции, то: DNn /Dt =B, или рождаемость; DNn/(NDt)=b, или рождаемость на единицу популяции. N может представлять всю популяцию или только ту ее часть, которая способна к размножению. Например, для высших организмов рождаемость обычно принято выражать в расчете на одну самку. В демографии рождаемость обычно выражают числом рождений на 1000 человек за год. Плодовитость определяют как среднее число оплодотворенных яиц, производимых за один цикл размножения или в течение всей жизни. Специфическую рождаемость можно определить как рождаемость, специфичную для разных возрастных групп популяции, или возрастную рождаемость. Максимальная рождаемость - это теоретически возможный верхний предел, которого популяция или способная к размножению часть популяции могла бы достигнуть при идеальных условиях. Трудности, связанные с определением этого термина очевидны, но он представляет интерес по двум причинам. 1. Максимальная рождаемость служит критерием для сопоставления с реализуемой рождаемостью. Так, утверждение, что рождаемость в популяции мышей составляет 6 детенышей на самку в год, будет иметь больше смысла, если известен предел, до которого она могла бы увеличиться в менее лимитирующих условиях. 2. Являясь постоянной величиной, максимальная рождаемость может быть использована при выведении уравнений для определения и прогнозирования скоростей увеличения популяции. Поскольку ценность понятия величины максимальной рождаемости состоит в том, что можно сравнивать с ней как с константой различные наблюдаемые величины рождаемости, использовать ее имеет смысл лишь в том случае, если известны условия, при которых она определяется, отсутствует лимитирующее влияние физических факторов, и размеры популяции, которые в свою очередь могут действовать как лимитирующий фактор, оптимальны. | |||||||||