молочное скотоводство. Молочное коневодство предусматривает получение кобыльего молока на специальных фермах и производство из него высокоценных пищевых, диетических и лечебных продуктов, прежде всего кумыса
 Скачать 3.26 Mb. Скачать 3.26 Mb.
|

|
Кислотность Кислотность кобыльего молока составляет 5°Т. Один градус Тернера составляет 0,009 процентов молочной кислоты. Титруемая кислотность молока обуславливается наличием белков, кислых солей и растворенного диоксида углерода. По мере хранения сырого молока титруемая кислотностью повышается вследствие развития молочнокислых бактерий. По своей кислотности (5°Т) молоко стоит ближе к женскому (4-5°Т) и намного ниже коровьего (16-18°Т), что обусловливается специфичностью белкового состава кобыльего молока. Бактерицидные свойства Бактерицидные свойства молока кобыл достаточно высокие. Молоко не содержит фермента пероксидазы; каталазное число равно 0,53-0,57, хлор–сахарное – 0,576. Бактерицидная фаза молока кобыл при 37°С продолжается 2½-4½ часа, а при пониженных температурах – более продолжительное время. Химический состав молока кобыл изменяется в зависимости от породы, кормления, лактации, кратности доения. Состав молока существенно изменяется в течение лактации, причем наибольшим изменениям подвержен белок: в первых порциях молозива его количество достигает 32%, но уже через 12 ч после выжеребки, по данным И.А. Сайгина, количество белка снижается до 4-4,3 %, а еще через 12 ч – до 3,4-3,6 %, удерживаясь на таком уровне примерно до 5-го дня. Содержание белка на уровне 3% сохраняется до 10-15 дня, а затем снижается до оптимального уровня. Примерно через час после выжеребки кобыл содержание лактозы в молозиве невелико – 2,9-3,4 % и лишь к 15-20 дню доходит до 6%, максимальное ее количество (7%) наблюдается через 1-3 месяца после выжеребки. На протяжении лактации количество жира в молоке сильно варьирует. Так, через час после выжеребки, количество жира в молозиве кобыл изменяется от 1 до 6%. В первые сутки молозиво кобыл темного цвета с зеленоватым оттенком. Кислотность его колеблется от 20 до 98°Т, лишь на 4-5-й день молоко приобретает нормальный белый цвет и свойственные ему вкус и запах (кислотность 6-8°Т). Строение вымени кобыл Вымя кобылы по анатомическому строению, а в связи с этим и по физиологии отдачи молока, существенно отличается от вымени коров. Для выработки правильной техники ручного и машинного доения, контрольного определения молочности, внедрения рационального распорядка дня на кумысных фермах, необходимо знать специфику строения и функции вымени кобыл. Молочная железа (вымя) у кобыл расположена в паховой области, масса вымени 1600 г, емкость – от 1 до 3 л. Это компактное образование округло-овальной формы, состоящее из двух половин – правой и левой. Размеры его сравнительно небольшие: окружность у основания 45-60 см, высота 10-15 см, длина сосков 3-5 см. Соски имеют коническую форму, сплюснутую у основания. Окружность сосков у основания 9-14 см. Форму и размер сосков нужно учитывать при конструировании доильных аппаратов для машинного доения. Каждая половина вымени в свою очередь, состоит из двух долей, имеющих один общий сосок, в котором заложены две сосковые цистерны, два сосковых канала и два выводных отверстия с запирательным сфинктерами. Каждая доля вымени состоит из огромного количества альвеол, которые снаружи покрыты миоэпителием. Мельчайшие выводные протоки альвеол, сливаясь между собой, образуют систему более крупных протоков. В каждую молочную цистерну открывается 10-20 крупных и множество мелких протоков. Между половинами вымени находится соединительнотканная перегородка, которая играет роль поддерживающей связки. Между собой половины вымени связаны нервами и кровеносными сосудами, но их молочные ходы и железистая ткань друг с другом не сообщаются. В пределах каждой половины вымени имеются две обособленные доли - передняя и задняя. Молочные ходы этих долей также не сообщаются. Каждая доля имеет молочную цистерну и сосковый канал с самостоятельным выводным отверстием, поэтому на каждом соске у кобылы по два отверстия – от передней и задней долей вымени. Передняя и задняя доли могут быть одинаковыми, но встречаются кобылы, у которых значительно больше развиты передние доли. Изредка встречаются кобылы с тремя долями в каждой половине вымени, которые развиты слабо и являются нежелательными, так как меняют требования к режиму доения. М 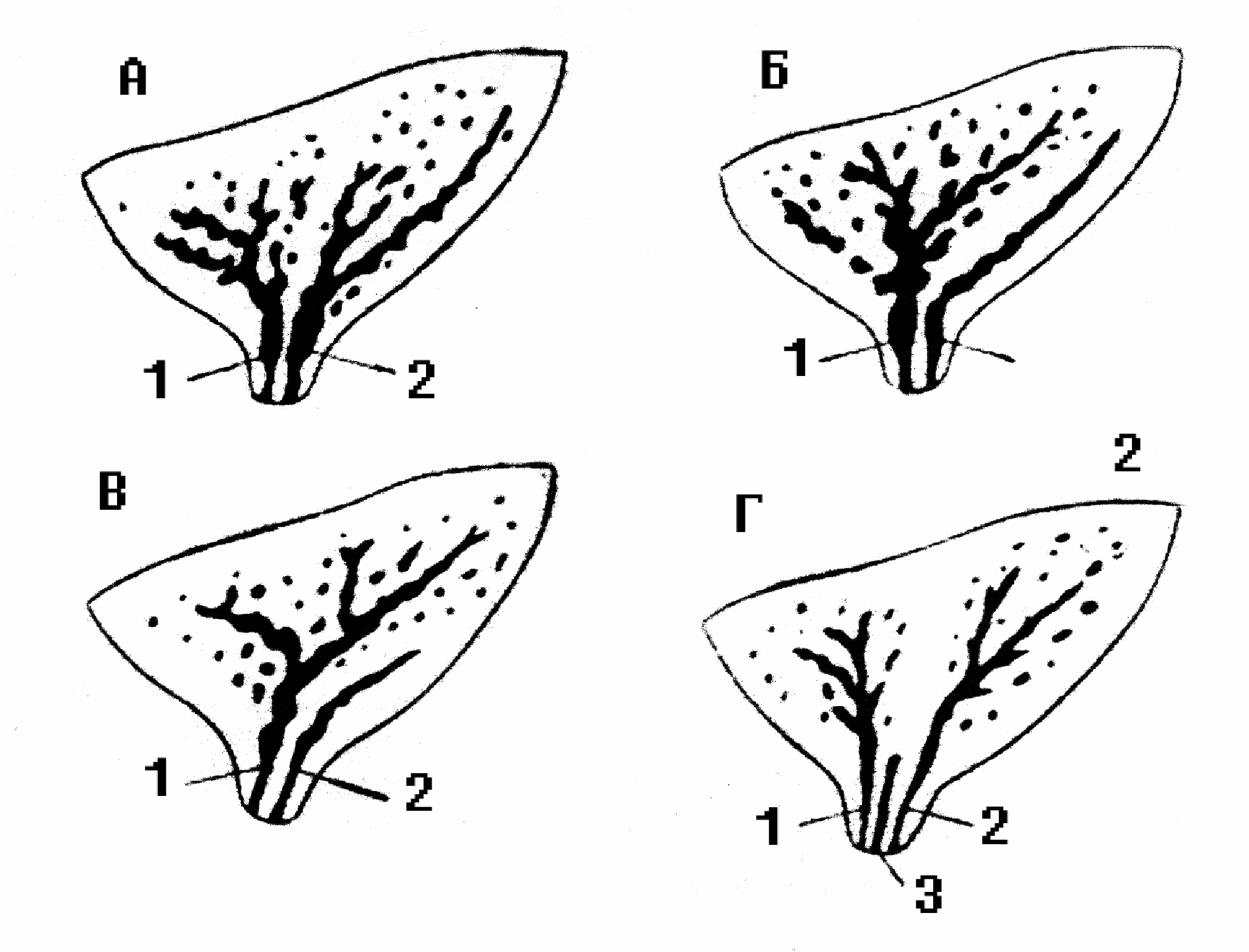 олочные цистерны в вымени отличаются небольшими размерами, в них может накапливаться лишь незначительная часть молока. Из цистерны вглубь железы ведут разветвляющиеся молочные ходы, имеющие сравнительно большое сечение. По мере ветвления диаметр ходов уменьшается, а число их увеличивается. Самые тонкие ветви ходов, в виде узких канальцев, видимые только под микроскопом, заканчиваются альвеолами (пузырьками), которые выстланы железистыми эпителиальными клетками, продуцирующими молоко. Вокруг молочных ходов расположены пучки волокон гладкой мускулатуры, под воздействием сокращения которых происходит выдаивание молока в момент сосания жеребенка или доения. Общий объем молочных ходов в 9-10 раз превышает объем сосковой и надсосковой цистерн. Из-за отсутствия развитых цистерн, образующееся в вымени кобылы молоко быстро заполняет крупные молочные ходы, а затем мелкие канальцы и альвеолы, где нарастает избыточное давление тормозящее дальнейший процесс молокообразования. Из-за особенности строения вымени кобылы необходимо часто сдаивать молоко. Вымя кобылы невелико по размерам, но богато железистой тканью и способно выделять столько же молока, сколько продуцирует вымя коровы, которое по размерам в несколько раз больше. Размеры вымени зависят от молочной продуктивности животного. На формирование емкостной функции вымени большое влияние оказывает возраст животного и число лактации. У молодых и старых кобыл емкость вымени меньше, чем у кобыл среднего возраста. Сосок у кобыл конической формы, несколько сдавленный с боков, стенка соска кобылы тоньше, чем у коровы. Кожный покров вымени гладкий, с нежными волосками, потовыми и сальными железами. На рост и развитие молочной железы оказывают влияние возраст, кормление и содержание животного, половое созревание, беременность, выжеребка, массаж вымени и др. При опорожнении вымени сначала выдаивают (или высасывает жеребенок) только небольшую порцию молока из цистерн. Именно из-за такой особенности строения вымени кобылы, принудительно выдоить руками или машиной можно лишь около 10% молока, то есть только то, которое находится в цистернах, 90% молока кобыла должна «отдать». В обычных условиях молокоотдача происходит под влиянием жеребенка, его вида, акта сосания, но можно у кобыл постепенно выработать условный рефлекс молокоотдачи на какой-нибудь раздражитель, например свисток или звук ритмичных ударов о подойник. Процесс молокоотдачи находится под контролем центральной нервной системы, по этой причине кобылы хорошо отдают молоко только при доении в привычной обстановке, а посторонний шум, присутствие новых людей, изменение распорядка работы могут вызвать торможение отдачи молока, вплоть до полного нарушения рефлекса молокоотдачи. На рисунке показаны рентгенограммы веточек артерий равных кусочков вымени кобылы и коровы, видна большая насыщенность кровеносными сосудами вымени кобылы. Это свидетельствует об обильном кровоснабжении, обеспечивающим высокую интенсивность образования молока. Как отмечалось выше, при опорожнении вымени вначале выдаивают (или высасывает жеребенок) только небольшую первую порцию молока, находящегося в цистернах. Затем молоко перестает выделяться, наступает пауза, так как для активизации гладкой мускулатуры, окружающей молочные ходы, нужно определенное время, примерно 10-40 секунд. Как только гладкая мускулатура вымени начнет сокращаться, молоко почти одновременно поступает из всех ходов и сразу в большом количестве наполняет цистерны. Если в это время задержать Р 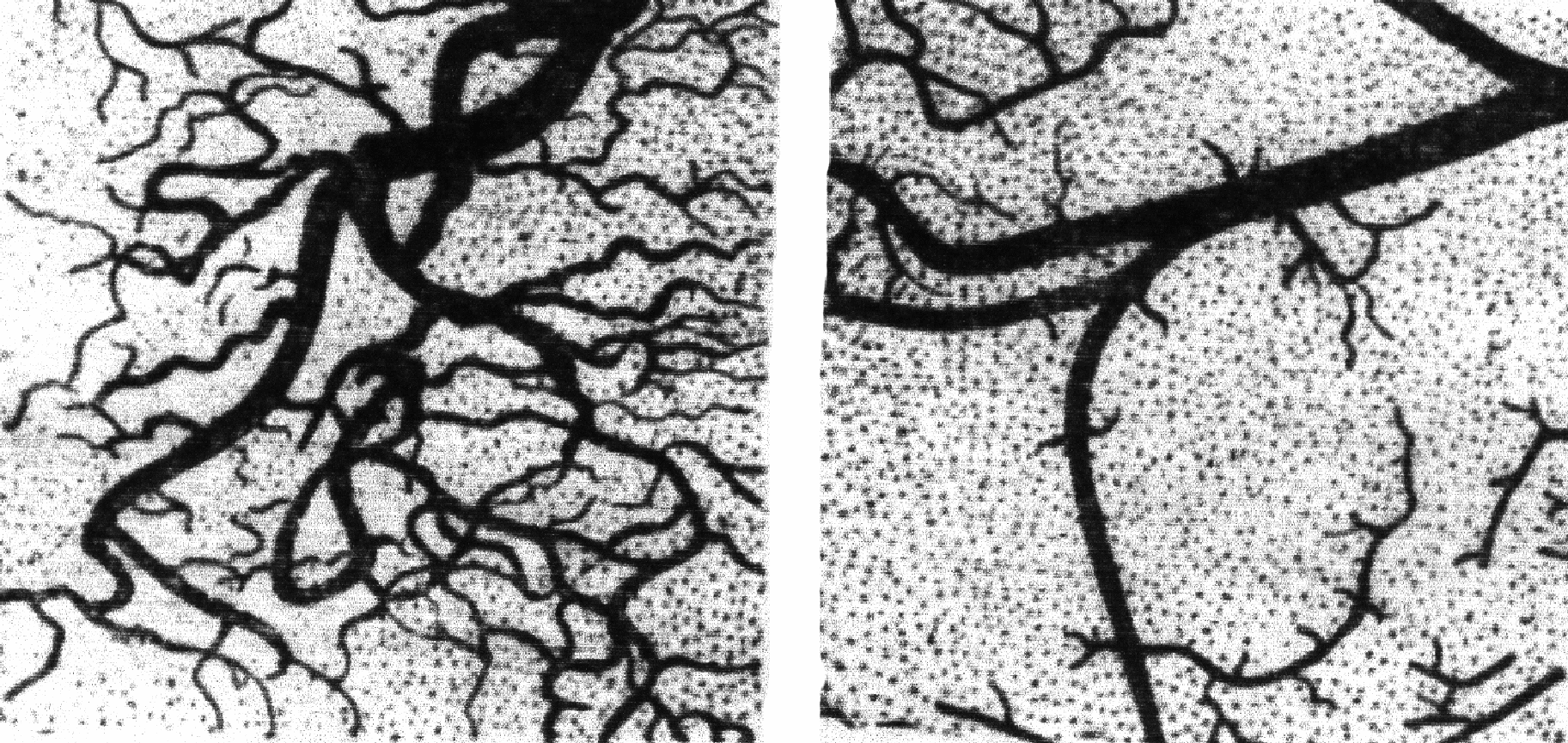 ис. опорожнение цистерн, затянуть доение, то сокращение гладкой мускулатуры прекратится и значительная часть молока останется в вымени. Чтобы избежать такого нежелательного явления, отдаивание основной (второй) порции молока надо проводить очень интенсивно, с расчетом полного опорожнения вымени за одну - полторы минуты (50-100 секунд). Из-за такой особенности молокоотдачи необходимо часто сжимать и разжимать пальцы рук или увеличить пульсацию доильной машины до 120-170 пульсаций в одну минуту. Особенности строения молочной железы кобылы Строение молочной железы как сложноорганизованного органа, состоящего из ряда тканей (железистой, мышечной, соединительной, жировой) обеспечивает оптимальные условия для образования специфического секрета. Железистая, или секреторная ткань у всех млекопитающих имеет вид гроздевидной пористой массы, состоящей из ветвящихся протоков, заканчивающихся альвеолярными расширениями. Каждая альвеола представляет собой небольшой пузырек (диаметр от 0.1 до 0,3 мм), стенка которого внутри выстлана одним слоем железистых (секреторных) клеток, а на наружной располагаются миоэпителиальные клетки, обладающие сократительной способностью. Слепой конец альвеолы имеет округлую форму: на противоположной стороне сужение переходит в выводной проток. Молочные альвеолы объединяются в отдельные группы, или дольки, имеющие длину около 1,5 мм, ширину 1 мм и высоту 0,5 мм: их объем 0,7-0,8 мм3. Тонкие выводные протоки, начинаясь от альвеол, постепенно сливаются с аналогичными протоками соседних альвеол во внутридольковые протоки. Дольки, в свою очередь, объединяются в более крупные образования – доли, имеющие широкие выводные протоки: диаметр долей достигает 2-17 мм. Крупные выводные протоки существенно отличаются по строению от начальных выводных протоков: слизистая оболочка их выстлана двухслойным цилиндрическим эпителием, а внутренняя стенка имеет складчатую поверхность. За эпителиальным слоем в стенке протока располагаются циркулярные и продольные гладкомышечные волокна. Устья протоков заканчиваются утолщением циркулярного слоя мышц в виде сфинктеров. Молоко, образующееся в альвеолах, по протоковой системе попадает в молочную цистерну-резервуар, который по своему анатомическому строению можно разделить на цистериальный отдел железы и цистерну соска. У кобылы на один сосок приходится две цистерны. Полость молочной цистерны отделена от цистерны соска круговой сфинктерообразной складкой. Соединительная ткань, или строма, молочной железы выполняет механическую, поддерживающую, защитную функцию. В соединительнотканной основе проходят кровеносные, лимфатические сосуды, нервы и она является продолжением соединительнотканного аппарата брюшной стенки. Брюшная фасция формирует соединительнотканную пластинку, которая, переходя в область основания вымени, образует подвешивающую связку, состоящую из двух листков, и делит вымя на правую и левую половины. Снаружи вымя поддерживается наружной фасцией, представляющей собой продолжение поверхностной брюшной фасции. Соединительная ткань формирует многочисленные перегородки между долями, дольками и альвеолами. Благодаря оптимальному соотношению эластических и каллогеновых волокон в строме при наполнении вымя больше растягивается в горизонтальном направлении (в стороны), чем в вертикальном (вниз). В иннервации молочной железы кобылы участвуют нервы пояснично-крестцового сплетения: в основном это наружный срамной нерв, в меньшей степени – подвздошно–паховые нервы. Молочная железа обладает развитым рецепторным аппаратом, благодаря которому осуществляется тактильная и температурная рецепция: афферентные нервы обеспечивают передачу соответствующей информации в различные отделы ЦНС. Афферентные нервы (асимптотический отдел вегетативной нервной системы) осуществляют как адренергическое, так и холинергическое неровное влияние на течение процессов развития и функционирования молочной железы. Кровоснабжение молочной железы у кобылы осуществляется следующим образом: кровь поступает, в основном, через наружную срамную артерию, а оттекает по наружной срамной и подкожной брюшной венам. Лимфатическая система молочной железы наряду с венозной, является системой оттока и состоит из замкнутых лимфатических капилляров и объединяющих их лимфатических сосудов. Лимфа оттекает по лимфатическим капиллярам и сосудам, проходит через лимфатические узлы и по выносящим лимфатическим сосудам поступает в поясничную лимфатическую цистерну, а из нее по грудному лимфатическому протоку попадает в переднюю полую вену и вместе с кровью в сердце. Физиология молокообразования и молокоотделения Процесс молокообразования Процесс секреции молока – это целостная реакция всего организма и молочной железы, протекающая циклично при участии нервной, кровеносной, эндокринной и пищеварительной систем. Молоко образуется в эпителиальных клетках альвеол и эпителии протоков из составных частей крови при участии ферментов и гормонов. По мере образования молоко из железистого эпителия выделяется в полость альвеол, накапливается в них, а потом в процессе доения поступает в протоки и молочную цистерну. Каждая эпителиальная клетка образует молоко со всеми присущими ему свойствами. В эпителиальных клетках синтезируются: молочный жир, лактоза, белки (казеины, β–глобулин). Витамины, минеральные вещества и некоторые белковые соединения (иммунные глобулины, сывороточный глобулин) переходят из крови в молоко без изменений. Установлено, что компоненты молока образуются из тех веществ, которые поступают с кровью в железу в виде его предшественников: молочный сахар – из глюкозы и галактозы; белок – из аминокислот; жир – из жирных кислот и глицерина, образующихся из жиров, белков и углеводов корма. Сущность процесса молокообразования заключается в поглощении из крови клетками железистого эпителия предшественников молока (аминокислот, липидов и др.), а затем в их синтезе и выделении (экструзии) из клетки в полость альвеолы в виде готового секрета. При переходе молока из клеток в альвеолы оно еще не окончательно синтезировано. Под влиянием ферментов и гормонов молоко дозревает в полости альвеол, причем часть основных элементов молока подвергается ферментативному расщеплению и всасывается обратно в кровь (реабсорбция), что в свою очередь стимулирует дальнейшую его продукцию. Реабсорбция и секреция взаимно обуславливают друг друга: нарушение реабсорбционных процессов влечет за собой и нарушение секреторных процессов образования молока. Существует несколько типов секреции молока. Основной тип секреции – мерокриновый. Эпителиальная клетка, получая кровь из сосудистого капилляра, выбирает из него вещества, которые используются для образования молока. Образующиеся капельки жира продвигаются в апикальную часть клетки и отсюда постепенно просачиваются через мембрану, оставляя на своем месте быстро исчезающую вакуоль. Клетка остается неповрежденной. Этот тип отмечают в разгаре лактации. В молозивный период секреция молока происходит по апокриновому типу, то есть происходит превращение дистального участка клетки в секрет. Апикальная часть клетки «выбрасывает» в просвет альвеолы вместе с секретом фрагменты цитоплазмы. В стадии инволюции происходит преобразование всей клетки в секрет молока, или голокриновый тип секреции. Структурная организация секреторного процесса Секретообразование тесно связано с функционированием секреторных клеток альвеолярного питания и их структурных компонентов. Наиболее важные звенья в цепи клеточных процессов, обеспечивающих образование и выделение из клетки секреторного продукта, следующие: а) синтез различных типов РНК (рРНК, мРНК, тРНК), обусловленный генетической информацией, содержащейся в ядре; б) участие мРНК и тРНК в синтезе белка на рибосомах (полисомах); в) деятельность структур эндоплазматического ретикулума в сегрегации и перемещении секреторного продукта; г) концентрация и «упаковка» секреторного продукта структурами комплекса Гольджи. Понятие «организация секреторного процесса» включает строгую регламентацию секретообразования. Многообразные процессы синтеза и переформирования первичных и промежуточных форм секреторного продукта строго локализованы и проходят в определенные промежутки времени. Эта пространственно-временная последовательность обязательно укладывается в рамки секреторного цикла. Секреторный цикл – это последовательное прохождение клеткой стадий метаболических и структурных перестроек, обеспечивающих полное превращение секреторного продукта от поступления веществ - предшественников до вывода из тела клетки готового сформированного продукта секреции. В ходе секреторного цикла на полисомах, расположенных на поверхности каналов эндоплазматического ретикулума, происходит процесс синтеза пептидных цепей. В полисоме на одну молекулу мРНК приходится несколько рибосом: образование полипептидных комплексов обеспечивает наиболее эффективное использование мРНК. Пройдя через одну и ту же серию кодонов мРНК, рибосомы синтезируют однотипный белок. На рибосомах синтез белка осуществляется за 2-3 минуты, в течение последующих 10 минут меченые аминокислоты обнаруживаются в составе пептидной или белковой молекулы в просвете каналов эндоплазматического ретикулума. В клетках молочной железы эндоплазматический ретикулум участвует и в синтезе молочного жира. Затем секреторный продукт поступает в комплекс Гольджи, где полипептидные цепи формируют мицеллы казеина за счет образования фосфатных и кальциевых связей. Структуры комплекса Гольджи осуществляют еще одну функцию секреторной клетки – синтез молочного сахара (лактозы). Лактоза, сформированная за время прохождения компонентов комплекса Гольджи, по цитоплазме вместе с гранулами казеина переходит в полость альвеолы при опорожнении везикул. Очевидно, это наиболее удачный путь выхода молочного сахара из клетки, поскольку лактоза неспособна проникать через клеточную мембрану. Электронномикроскопическое исследование секреторных клеток позволяет детализировать процесс удаления секрета. Крупная жировая капля (глобула) приближается к апикальной мембране и облекается ею. По мере увеличения контакта спазматической мембраной капля все больше выступает просвет альвеолы. Огибая жировую каплю, плазматическая мембрана сужается у основания и перешнуровывается, а затем отрывается и попадает в полость альвеолы. Выведение из клетки белка и лактозы происходит иначе: заключенные в мембраны конденсирующие вакуоли подходят к апикальной мембране, мембрана вакуоли соединяется с плазматической мембраной, затем полость вакуоли полностью раскрывается и ее содержимое опорожняется в просвет альвеолы. Альвеола – структурно-функциональная единица молочной железы. Как показали исследования на культивированных изолированных клетках молочной железы, уровень их секреции оказывается значительно ниже, чем в клеточном сообществе. То есть для сохранения секреторной функции железистых органов и, в частности молочной железы, необходима целостность клеточного комплекса как функциональной единицы. Взаимодействие между миоэпителиальными и секреторными клетками возникает в ходе функционального сопряжения процессов выведения ранее синтезированного молока и включения клеток альвеолярного эпителия в новый секреторный цикл. Выведение молока из полости альвеол и возбуждение секреторного процесса в железистых клетках воспроизводится гормоном окситоцином и медиатором нервной системы – ацетилхолином. Кроме того, при блокаде М-холирецепторов реакция клеток альвеолярного эпителия не развивается, хотя сокращение эпитетелия и выведение молока из полости альвеолы происходит. В миоэпителиальных клетках на основании гистохимического исследования выявлена возможность синтеза ацетилхолина и обнаружена активность специфического фермента ацетилхолитрасферазы. Система инактивации ацетилхолина представлена в молочной железе холиэстеридами, среди которых обнаружены как ацетлхолинэстераза, так и ацилгидролазы. Действие антихолиэстерадных препаратов, предупреждающих гидролиз ацетилхолина, обеспечивает потенцирующий эффект. Таким образом, можно говорить о существовании своеобразного механизма передачи регулирующих влияний с сократительной структуры альвеолы (миоэпителиальной клетки) на секреторные клетки с помощью медиатора (ацетилхолина), который синтезируется в миоэпителии и выделяется в пространство между секреторной и миоэпителиальной клетками и улавливается холирецепторами на мембране секреторной клетки. Сокращение миоэпителиальных клеток развивается при действии окситоцина и ацилихолина, причем клетки обладают самостоятельными окситоциновыми и холирецепторами. Сократительный аппарат миоэпителиальных клеток аналогичен таковому у клеток гладкой мускулатуры и представлен миофибриннами, заполняющими тело и отростки клеток. Кровоснабжение альвеол существенно меняется в ходе секреторного цикла, причем расширение микрососудов и раскрытие дополнительных капилляров обеспечивают рабочую гиперемию органа. Регистрация кровенаполнения сосудов, окружающих альвеолу, позволяет обнаружить наличие двухфазной реакции кратковременной вазоконструкции (сужение сосудов), обусловленной действием окситоцина (структурного аналога вазопрессина) и длительной вазодилатацией (расширением сосудов). Интенсивность секретообразования тесно связана с полиацепным кровоснабжением органа: объемный кровоток значительно возрастает при становлении лактации и увеличении молочной продуктивности в лактационный период. |