|
|
Пародонтология. Необходимо помнить, что ответственность за дозирование или введение лекарственного средства несёт врачстоматолог
§
ю
(В ГЛ
m О)
ш
та
С
го
Рис. 3.2. Метаболиты арахидоновой кислоты играют важную роль в развитии воспалительной реакции при хронических заболеваниях. При нормальных условиях арахидоновая кислота, связанная с фосфолипидами, создаёт эстры мембраны клетки. Вследствие разрушения тканей активируются фосфолипазы и высвобождается арахидоновая кислота (глкжокортикоиды препятствуют активации фосфолипазы). Проста гландины, простациклин и тромбоксан образуются вследствие активации циклооксигеназы СОХ1 и СОХ2 (торможение нестероидными противовоспалительными средствами), в то время каклипооксигеназа способствует формированию леикотриенов. Большое количество метаболитов арахидоновой кислоты проявляют высокую биологическую активность (табл. 3.1).
Таблица 3.1 Биологические функции метаболитов арахидоновой кислоты
Функции
М етаболит етаболит
 Простагландины Вазодилатация Возникновение боли Агрегация тромбоцитов Влияние на Т-лимфоциты, ингибирование продукции цитокинов Стимулирование остеокластов Лейкотриены Повышают сосудистую проницаемость Способствуют адгезии PMN к эндотелию Стимулируют хемотаксис PMN и эозинофильных гранулоцитов Индуцируют липосомную дегрануляцию PMN Повышают рост Т-лимфоцитарного хемокинеза
Продукт реакции - энзиматический катализатор следующего этапа.
Активация комплемента происходит:
Классическим путём посредством комплексов антиген/антитело.
Альтернативным путём посредством полисахаридов микроорганизмов (рис. 3.3).
Центральный компонент - это протеин СЗ с молекулярной массой 195 килодальтон и
концентрацией плазмы 1,2 мг/мл:
- При нормальных условиях больший фрагмент СЗЬ, спонтанно очень медленно возни кающий при наличии Мд 2+, может создать комплекс с компонентом комплемента - фак-
Патогенез гингивита, вызванного зубной бляшкой
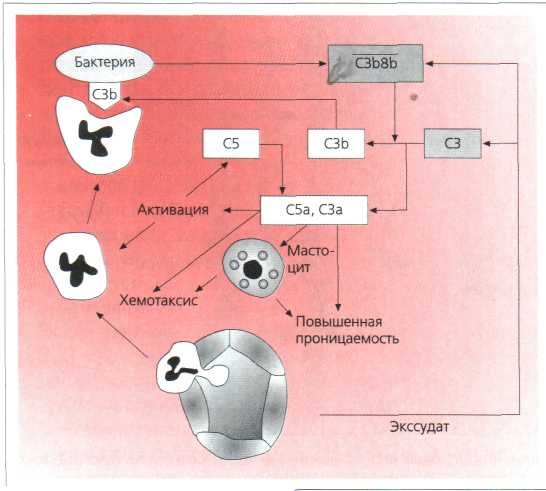 Рис. 3.3. Острая воспалительная реакция вследствие активации альтернативного комплементного пути. СЗ-конвертаза СЗЬ8Ь стабилизирована на поверхности бактерий и расщепляет большое количество СЗ на фрагменты СЗа и СЗЬ. СЗЬ связывается с бактерией, СЗа активирует С5. Продукт расщепления С5а является хемотаксиче-ским фактором для PMN, которые покидают сосуды и управляют бактерией. СЗа и С5а это анафилатоксины, побуждающие дегрануляцию тучных клеток (мастоцитов). Тучные клетки высвобождают вазоактивные медиаторы, способствующие насыщению ткани PMN и комплементом. Рис. 3.3. Острая воспалительная реакция вследствие активации альтернативного комплементного пути. СЗ-конвертаза СЗЬ8Ь стабилизирована на поверхности бактерий и расщепляет большое количество СЗ на фрагменты СЗа и СЗЬ. СЗЬ связывается с бактерией, СЗа активирует С5. Продукт расщепления С5а является хемотаксиче-ским фактором для PMN, которые покидают сосуды и управляют бактерией. СЗа и С5а это анафилатоксины, побуждающие дегрануляцию тучных клеток (мастоцитов). Тучные клетки высвобождают вазоактивные медиаторы, способствующие насыщению ткани PMN и комплементом.
тором В. После расщепления энзимом фактора D образуется СЗЬВЬ с энзиматиче-ской активностью (СЗ-конвертаза).
К онвертаза СЗ (СЗЬВЬ) стабилизируется на поверхности бактерии благодаря белку онвертаза СЗ (СЗЬВЬ) стабилизируется на поверхности бактерии благодаря белку
сыворотки пропердина и активируется полисахаридами микроорганизмов.
Конвертаза СЗ расщепляет большое количество СЗ на фрагменты СЗа и СЗЬ.
СЗЬ фиксирована к бактерии.
СЗа активирует С5, которая расщепляется на С5а и С5Ь.
Компоненты комплемента СЗа и С5а проявляют биологические защитные функции:
С5а - значимый хемотактический фактор для ПМЯ (PMN).
СЗа и С5а являются анафилотоксинами. Они вызывают:
Дальнейшая активация системы комплемента влечет за собой образование пор в клеточной мембране с лизисом бактерий:
С5Ь свободно связывается с СЗЬ.
С6 и С7 последовательно связывается с С5Ь.
С8а и С9 образуют комплекс, атакующий мембрану.
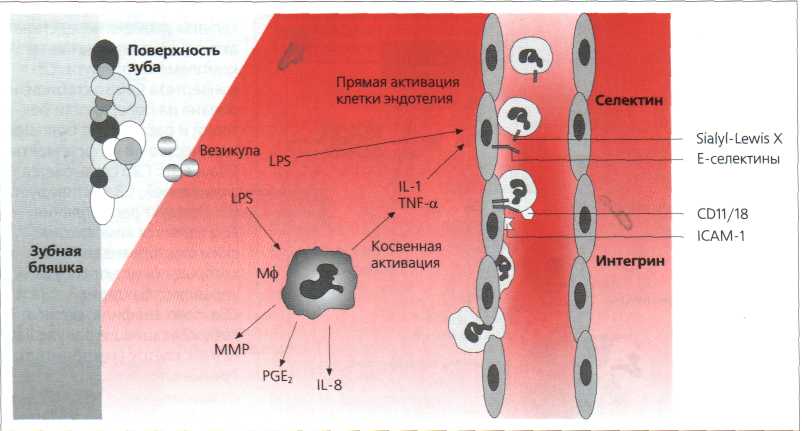
Для того, чтобы лейкоциты покинули сосудистое русло, необходимо усилить адгезив-ность молекул в клетках эндотелия и лейкоцитах (рис. 3.4):
Бактериальные продукты как липополисахариды (LPS) грамотрицательных бактерий
или протеины в форме расщеплённых фрагментов бактериальной поверхности могут
вступать в прямые реакции с клетками эндотелия капилляров и сосудов в десневой
соединительной ткани.
Возможна также непрямая активация посредством макрофагов, которые после кон-
S
ё
>s о
го
X
л
X I
го со 2
I
го
і
a>
§
ю го
І
s
го
00
Патогенез гингивита, вызванного зубной бляшкой
 s х го со ф s х го со ф
§
ю та гч
сі
си
си
I та
Рис. 3.4. Активация клеток эндоделия происходит непосредственно через расщеплённую оболочку липополисахаридов (LPS) грамотрицательных бактерий на поверхности зуба или косвенно посредством провоспалительные цитокинов: интерлейкин 1 (IL-1) и фактор альфа некроза опухоли (TNF-α), высвобождаемых макрофагами (Мф) наряду с другими цитокинами и металлопротеиназами матрикса (ММР), простагландином Е2 (PGE2) и хемокином IL-8. Е-селектин активируется на внутренней стенке сосудов. Посредством Lewis-антигенов групп крови он свободно связывается с гранулоцитами, скорость которых в кровотоке значительно уменьшается. Когда нейтрофильные гранулоциты перемещаются по эндотелию, то может произойти прочное сцепление адгезивного рецептора лейкоцита CD11/18 и межклеточной адгезивной молекулы ІСАМ-1. Затем гранулоцит мигрирует за пределы сосуда (по Darveau et al., 1997).
такта с LPS или протеинами бактерий высвобождают провоспалительные цитокины интерлейкин 1 (IL-1) и фактор альфа некроза опухоли (TNF-α).
В обоих случаях на внутренней стороне сосуда активируется адгезивная молекула - се-
лектин Е, которая принуждает гранулоциты перемещаться по эндотелию.
После контакта адгезивного лейкоцитарного рецептора CD 11/18 с эндотелиальным
интегрином 1C AM-1 (внутриклеточная адгезивная молекула) возникает амёбовидный
диапедез (рис. 3.4).
> Миграция гранулоцитов связана с градиентами хемотаксических факторов:
Бактериальные пептиды, напр. N-Formyl-methionyl-leucyl-phenylalanin (FMLP).
Хемотаксический цитокин (хемокин) 1L-8; последний в большом количестве пере
даётся клетками эпителия в соединительную ткань.
Молекулы, высвобождаемые другими гранулоцитами, напр, лейкотриен В4
Компоненты комплемента С 5а.
Для целенаправленной миграции гранулоцитов необходимо достаточное количество
соответствующих функциональных поверхностных рецепторов.
Соединительный эпителий активирует интегрин 1C AM-1:
Это способствует адгезии нейтрофильных гранулоцитов и некоторых лимфоцитов
посредством адгезивных лейкоцитарных рецепторов CD11/18.
Таким образом соединительный эпителий сигнализирует о наличии бактерий рас
положенной под ним соединительной ткани.
 Патогенез гингивита, вызванного зубной бляшкой *t Патогенез гингивита, вызванного зубной бляшкой *t
В соединительном эпителии, а впоследствии также в эпителии кармана, имеются нерв- '5
ные элементы. Кроме своей барьерной функции соединительный эпителий выполняет *
сенсорную и сигнализирующую функции.|
Нейтрофильные гранулоциты достигают борозды; при поддержке комплемента и v§
антител (опсонин) происходит фагоцитоз бактерий и их продуктов. >s
Лейкоциты связываются с опсонизованными бактериями посредством рецепторов Fc: і
FcyRI, проявляя высокую степень сходства, связывается с гаммаглобулинами под- >»
классов IgG 1, IgG3 и IgG4. x
FcyRI 1 и FcyRIII со слабой степенью сходства связываются с комплексом антител или 3
агрегациями из IgG 1 и IgG3. і
FcyRH как единичный Fc-рецептор связывается с lgG2 подкласса IgG, который фор- м
мируется главным образом взамен капсульных антигенов полисахаридов грамотри- 3
цательных бактерий. ш.
> После фагоцитоза фагосомы сплавляются с цитоплазматическими гранулами (табл. н
3.2), превращаясь в фаголизосомы. Бактерии уничтожаются внутри клетки. Этот про- о
цес осуществляется несколькими путями: о
- Кислородозависимое уничтожение.Jo"
. анион супероксида: NADPH + 2 О2^^S^ NADP + Н+ + 2 О2" ,=
+ 2 Н2О -»2 Н2О2 -> *ОН s
миелопероксидазосодержащий компонент: гипохлорная кислота, хлорамин го
- Кислородонезависимое уничтожение:S
миелопероксидаза о
дефензины: специфические антитиотические пептиды *го
катепсины Л5
лактоферрин §2
лизоцим. 2!
> Внеклеточное вьщеление содержимого гранул с целью защиты инвазивных бактерий °
может вызвать значительное разрушение тканей: С
Эндопептидазы (протеазы) инактивируются посредством а2-макроглобулинов и аі -ин- т
гибиторов протеиназы.
С другой стороны, сверхактивные протеазы микроорганизмов пародонта могут рас-
Таблица 3.2 Цитоплазматические гранулы полиморфноядерных гранулоцитов
Энзимы Первичные (азурофильные) гранулы Вторичные (специфи-
ческие) гранулы
Бактерицидные энзимы Миелопероксидаза Лизоцим
Лизоцим
Нейтральные протеазы Эластаза Коллагеназа
Катепсин G Протеиназа 3
Кислые гидролазы Ы-ацетил-р-глюкосаминидазы
Катепсин В, D р-глюкуронидаза р- гл ицерофосфатаза р-маннозидаза
Другие Дефензин Лактоферрин
Катионные протеины Кобалофилин
Увеличение фактора бактерицидность/ CR3
проницаемость Цитохромы
Патогенез гингивита, вызванного зубной бляшкой
*s щепить эти ингибиторы. Пример: гингипаин, продуцируемый бактерией P. gingivalis.
* > Концентрация ряда протеинов плазмы, т.н. протеины острой фазы, значительно повы-
к шается под влиянием провоспалительных медиаторов таких как IL-1:
»§ - Протеины острой фазы со значительным увеличением
>s • С-реактивный протеин (CRP) - связывает комплемент, опсонизирует
і • маннозосвязывающий протеин - связывает комплемент, опсонизирует
>, • милоид сыворотки Р: инициирует отложение амилоида
х - Протеины острой фазы с умеренным увеличением (ингибирование бактериальных
| протеаз): \
і • аг-макроглобулин
со • си - ингибитор протеиназы.
Ранний гингивит
в
х > В течение 2-й и 3-й недель ненарушенной аккумуляции зубной бляшки наблюдаются
§; первые симптомы воспаления: гиперемия и отёчность. Эти симптомы возникают в ре-
§. зультате:
™ - нарушение микроциркуляции
>s - увеличения проницаемости сосудов с повышенным выделением протеинов плазмы.
> Клинически установлено значительное увеличение образования десневой жидкости.
ш > Раннее поражение - это проявление ответной иммунной реакции на антигены зубной
с- бляшки:
v§ - типичное поражение десны у здоровых детей и подростков
m - сходство с лимфоидными тканями.
3 > Соединительный эпителий содержит клетки, запускающие иммунные реакции:
J - специальные Т-клетки слизистой оболочки
о - клетки Лангерганса, представляющие антиген
и - дентритные клетки.
с > В соединительной ткани образуется инфильтрат преимущественно из Т-лимфоцитов,
составляющих 10-15% объёма неприкреплённой десны (рис. 3.5). >• В-лимфоциты встречаются только в незначительном количестве. Они превращаются в
клетки, продуцирующие антитела.
>Системный гуморальный иммунный ответ:
Антигенный бактериальный материал поглощается клетками Лангерганса в эпителии
и макрофагами в соединительной ткани десны и транспортируется в региональную
лимфоидную ткань.
Стимуляция лимфоцитов, вырабатывающая специфический иммунный ответ:
клетки плазмы продуцируют специфические антитела в лимфатических узлах
антитела через системы кровообращения попадают в десну
антитела можно обнаружить в десневом экссудате
> Остается неясным, вырабатывается ли местно специфический иммунный ответ:
Для пародонтальных тканей специфические В- и Т-клетки должны пролифериро-
вать в региональных лимфоузлах и поступать в кровоток.
Затем эти лимфоциты попадают в пародонт, где выполняют свои гуморальные и кле
точные функции.
Клетки плазмы, контролируемые ТЬ2-клетками, продуцируют антитела. Клеточный
иммунный ответ регулируется ТЫ -клетками (см. далее).
>Макрофаги всегда представляют только незначительную часть популяции клеток:
Патогенез гингивита, вызванного зубной бляшкой in
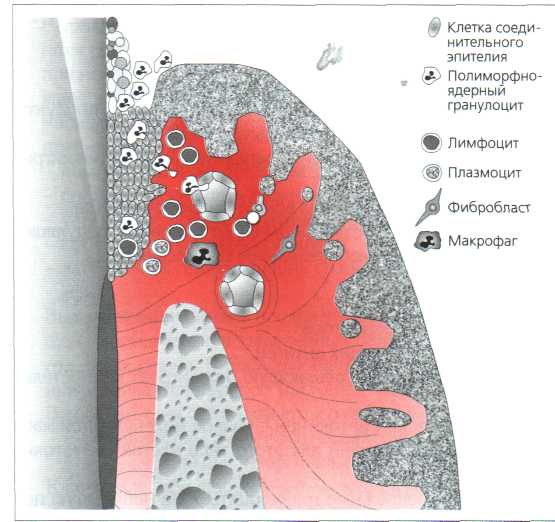
Р ис. 3.5. Характеристика раннего поражения. Дальнейшее увеличение проницаемости сосудов способствует интенсивному притоку протеинов плазмы, включая протеины острой фазы, комплемент и плазмин. PMN в большом количестве диффундируют в соединительную ткань и соединительный эпителий, что усиливает активирование хемотокина IL-8. Вследствие этого происходит разрыхление дна борозды и латеральной пролиферации базальных клеток. Активированные макрофаги продуцируют провоспалительные цитокины IL-1, IL-6, IL-10, TNF-α и такие хемотокины как IL-8 и МСР-1 (хемоатрактант протеин), а также PGE2 и коллагеназу тканей. Благодаря этому обеспечивается накопление Т-лимфоцитов и моноцитов из сосудистого русла. Т-лимфоциты взаимодействуют с фибробластами, подвергаемыми цитопатическим изменениям. Инфильтрат раннего поражения составляет 10-15% объёма неприкреплённой десны (адаптировано по Page & Schroeder, 1990). ис. 3.5. Характеристика раннего поражения. Дальнейшее увеличение проницаемости сосудов способствует интенсивному притоку протеинов плазмы, включая протеины острой фазы, комплемент и плазмин. PMN в большом количестве диффундируют в соединительную ткань и соединительный эпителий, что усиливает активирование хемотокина IL-8. Вследствие этого происходит разрыхление дна борозды и латеральной пролиферации базальных клеток. Активированные макрофаги продуцируют провоспалительные цитокины IL-1, IL-6, IL-10, TNF-α и такие хемотокины как IL-8 и МСР-1 (хемоатрактант протеин), а также PGE2 и коллагеназу тканей. Благодаря этому обеспечивается накопление Т-лимфоцитов и моноцитов из сосудистого русла. Т-лимфоциты взаимодействуют с фибробластами, подвергаемыми цитопатическим изменениям. Инфильтрат раннего поражения составляет 10-15% объёма неприкреплённой десны (адаптировано по Page & Schroeder, 1990).
3
ё
І
сі
*
і
m
З m
та
5
о. го
с
>5 S
та
S
§
ю
Я
го a>
ш
та
При экспозиции на LPS они превращаются в эффекторные клетки и выделяют:
провоспалительные цитокины
простагландин Ег
хемокины
металлопротеиназы матрикса как коллагеназу.
Благодаря этому обеспечивается постоянное пополнение лимфоцитов и моноцитов. Выраженный гингивит При дальнейшей аккумуляции бактериальной зубной бляшки через некоторое время у взрослых развиваются выраженные поражения. Воспалительный процесс вызывает образование десневого кармана. Механизм его развития связан с внутриэпителиальной щелью с последующей дистрофией клеток соединительного эпителия.
Потеря биологической связи между соединительным эпителием и поверхностью эмали.
Образование эпителия кармана:
Патогенез гингивита, вызванного зубной бляшкой
 S S
3
І
а. го
го
со <и
§ я
ГО
ш
S
пролиферация эпителиальных тяжей в инфильтрированную соединительную
ткань
микроизъязвления между Эпителиальными тяжами.
• наличие остатков соединительного эпителия на дне десневого кармана.
Вследствие пролиферации бактерий образуется поддесневая микрофлора, продукты
обмена которой оказывают прямое влияние на соединительную ткань.
Специфические, значительно отличающиеся популяции воспалительные клетки мигри
руют в соединительную ткань и в карман:
Нейтрофильные гранулоциты образуют плотный вал вокруг микроорганизмов.
Макрофаги, лимфоциты и плазматические клетки формируют основные популяции
клеток в соединительной ткани. Происходит избирательное перемещение:
антигенспецифических клеток памяти
активированных лимфоцитов
слизистьрс, положительных Т-клетокуб-рецептора
CD la-положительных клеток, представляющих антиген.
- В регулировании целенаправленной миграции решающую роль играет избиратель
ная экспрессия адгезивных молекул.
Особое значение придается хемотаксическим цитокинам с низкой молекулярной массой и потенциальными, характерными свойствами для этого типа клеток (хемотоки-ны):
- IL-8 реагирует специфически на нейтрофильные гранулоциты и на небольшую по
пуляцию лимфоцитов.
- МСР-1 (monocyte chemoattractant protein 1) отвечает за миграцию моноцитов.
Генерируемые плазматическими клетками антитела не обязательно направлены против
антигенов зубной бляшки. Микроорганизмы зубной бляшки скорее всего способству-
Р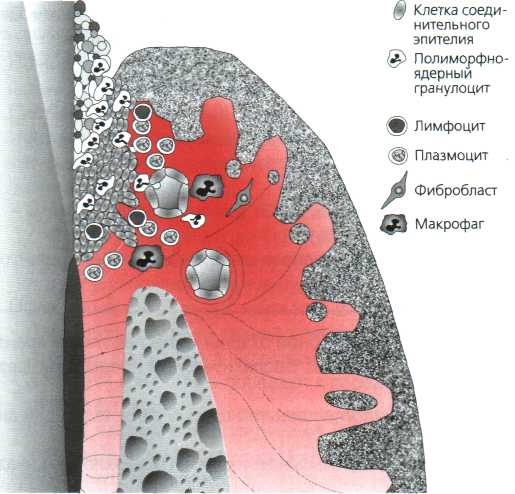 ис. 3.6. Характеристика выраженного поражения. Чётко выражена латеральная пролиферация соединительного эпителия. Через внутриэпителиальную щель образуются пародонтальные карманы с последующей пролиферацией в поддес-невой участок. Острый компонент воспаления пер-систирован. В инфильтрате преобладают теперь специфические популяции моно-нуклеарных клеток. КТ-лимфоцитам присоединяются В-лимфоциты и плазматические клетки, выделяющие гаммаглобулины, в основном неспецифические, поликлональные антитела (адаптировано по Page &Schroeder, 1990). ис. 3.6. Характеристика выраженного поражения. Чётко выражена латеральная пролиферация соединительного эпителия. Через внутриэпителиальную щель образуются пародонтальные карманы с последующей пролиферацией в поддес-невой участок. Острый компонент воспаления пер-систирован. В инфильтрате преобладают теперь специфические популяции моно-нуклеарных клеток. КТ-лимфоцитам присоединяются В-лимфоциты и плазматические клетки, выделяющие гаммаглобулины, в основном неспецифические, поликлональные антитела (адаптировано по Page &Schroeder, 1990).
Патогенез гингивита, вызванного зубной бляшкой
ют неспецифической, поликлональной стимуляции В-клеток. >s
Выраженный гингивит очень распространён. Практически у каждого взрослого име- *
ются соответствующие поражения. Неизвестно, по истечении какого периода времени к
при ненарушенной аккумуляции зубной бляшки возникают явные поражения. В насто- vo
ящее время предполагается продолжительность от нескольких недель до нескольких «s
месяцев. Патогистологически выраженный гингивит характеризуется следующими і
признаками (рис. 3.6): >,
персистирующие острые компоненты воспаления х
специфические популяции воспалительных клеток в инфильтрате 2
увеличение доли плазматических клеток х
последующая потеря коллагена в
латеральная пролиферация соединительного эпителия, образование десневого кар- 3
мана. • "І
Выраженный гингивит продолжительное время может оставаться стабильным. В об
щем имеется точно определённый баланс между бактериальным вызовом и иммунным о
ответом хозяина. В последующем после продолжительного неопределённого состоя
ния развивается прогрессирующее поражение (пародонтит). §■
В соответствии с этим имеются две формы выраженного гингивита: ^
- В большинстве случаев это независимое, самостоятельное и стабильное поражение, s
реже - первый шаг к маргинальному пародонтиту. <и
Клинически обе формы невозможно различить. Гистопатологически в активных пора- £
жениях вероятно имеется более высокая плотность плазматических клеток (более 50%). о
ю
я і
g
as
Патогенез маргинального пародонтита
■S Прогрессирующее поражение
ж
>Постоянная атака организма хозяина бактериями, заселяющими поддесневой участок,
по истечении неопределенного времени вызывает разрушение специфических и не-
J? специфических защитных механизмов:
о - Начальная потеря прикрепления взаимосвязана с увеличением в поддесневой флоре
ю В. Forsythus, С. rectus и Selemonas noxia.
f> - Активные фазы с клинической явной потерей прикрепления появляются редко и на
2 короткое время.
> При прогрессировании поражения и обнажения границы эмаль/цемент достигается
стадия маргинального пародонтита. Поражаются все структуры пародонта:
2 - Окончательное заселение поддесневой флоры зубной бляшки.
а - Пролиферация тонкого эпителия кармана со своеобразными эпителиальными тяжами
re возникает вместе с апикальной пролиферацией остатков соединительного эпителия
і на поверхности цемента, покрытого дегенерированными волокнами надальвеоляр-
ct ного волокнистого аппарата.
о. - В патологически изменённый цемент корня на участке пародонтального кармана
с бактерии проникают через резорбционные лакуны.
>s > При увеличении интенсивности воспаления снижается концентрация хемотокинов:
х - Спадает миграция нейтрофилов
2 - Активация процесса возможна только в соединительной ткани; при местном дефи-
с; ците (х2-макроглобулина и си -ингибитора протеиназы имеет место эксцессивная дест-
\о рукция пародонта.
m > Поверхностные антигены оказывают влияние на дифференциацию Т-лимфоцитов на:
3 - Т-клетки-помощники (Th: CD4+) связаны с антигеном МНС-П-комплекса и различают
J эпитопы, представляемые соответствующими клетками. Регулирование клеточного и
о гуморального иммунного ответа осуществляется посредством различных цитокинов
й (табл. 3.3):
ТЫ-клетки продуцируют интерферон гамма (INF-γ) и IL-2: клеточные реакции и
гиперчувствительность; стимуляция нейтрофильных гранулоцитов и активация ма
крофагов; продуцирование провоспалительных цитокинов, включая IL-1 и TNF-α.
ТЬ2-клетки продуцируют 1L-4, IL-5, IL-6, IL-10: влияние на аллергические реакции
и реакции на антитела; дифференциация В-клеток и плазмоцитов; стимуляция туч
ных клеток и эозинофильных гранулоцитов.
ThO- клетки продуцируют INF-γ, IL-2, IL-4 и IL-5.
- Цитотоксические/супрессорные Т-клетки (Тс, Ts: CD8+) связывают антигены
класса I МНС-комплекса.
Цитотоксические Тс-клетки продуцируют IL-10 и INF-γ.
Тэ-супрессорные клетки продуцируют IL-4.
> Соотношение CD4- и CDe-клеток в воспалительном инфильтрате повышается при мар
гинальном пародонтите. Среди клеток-помощников доминирующими, по-видимому,
являются Th2- клетки:
Их цитокины усиливают локальный гуморальный иммунный ответ.
В частности, IL-4 стимулируют В-лимфоциты к продукции IL-1.
Одновременно продукция 1L-1 подавляется макрофагами.
> Под влиянием LPS и провоспалительных цитокинов и медиаторов IL-1, TNF-α или PGE2
клетки соединительного эпителия, фибробласты и васкулярные эндотелиоциты разви
вают генетически запрограммированную, деструктивную активность:
Патогенез маргинального пародонтита
ІЛ
Таблица 3.3 Происхождение и функции некоторых цитокинов
Цитокины Происхождение
функции во время воспаления
IL-1
Макрофаги (Мер), фибробласты
Пролиферация активированных В- и Т-клеток
Индукция PGE2 и продукции цитокинов микрофагов
Экспрессия эндотелиальных адгезивных молекул
Индукция продукции IL-6, INF-pi и GM-CSF
Индукция лихорадки, протеинов острой фазы, активности
остеокластов
TNF-α
Мф, Т-клетки
Индукция протеинов острой фазы
Активация фагоцитов
Индукция INF-γ, TNF-α, IL-1, GM-CSF, IL-6
INF-γ
Т-клетки, естественные клетки-киллеры (NK, natural killer)
Т-клетки
Т-клетки, тучные клетки (лаброциты), базо-фильные гранулоциты
Индукция ТМ-клеток
Ингибирование активности IL-4
Усиление продукции IL-12
Стимуляция активности Мф, цитотоксических Т-клеток и NK
IL-2 IL-4
Стимуляция роста активированных Т- В-клеток, NK
Индукция дифференцирования Тп2-клеток
Ингибирование активности, индуцированной IL-2 и INF-γ
Ингибирование продукции IL-12
Индукция пролиферации и дифференциация Т-клеток
Индукция пролиферации Т-клеток
IL-5 IL-6 IL-10
Регулирование моноцитарной продукции IL-1, TNF-α и IL-6
Т-клетки, тучные клетки
Th-клетки, Мф, тучные клетки, фибробласты
Т- и В-клетки, моноциты и Мф
Пролиферация активированных В-клеток Продукция IgM и IgA
Рост и дифференцирование В- и Т-клеток Индукция протеинов острой фазы
Усиление ТЬ2-реакций при одновременном угнетении ТМ-реакций
Угнетение пролиферации и продукции цитокинов активированных Т-клеток
Угнетение моноцитарной продукции IL-1, IL-6, IL-8 Рост продукции IL-1ra Рост пролиферации и дифференцирования В-клеток
IL-12
В-клетки, Мф, дендритные клетки, кера-тиноциты,нейтро-фильные гранулоциты
Т-клетки
Ключевая роль при индукции ТМ-реакций Стимулирование роста и цитотоксической активности NK и Т-клеток
IL-13
Регулирование продукции IL-12
Синтез Тгі2-реакции
Стимулирование В-клеток
Ингибирование продукции цитокинов макрофагов
2 ё
1
X
го
00
І
та
та ш o>
|
|
|
 Скачать 5.4 Mb.
Скачать 5.4 Mb.