1. Первичная структура белков
 Скачать 1.7 Mb. Скачать 1.7 Mb.
|

Снижение значений.
Нуклеозиды— это гликозиламины, содержащие азотистое основание, связанное с сахаром (рибозой или дезоксирибозой). Нуклеозиды выполняют только метаболическую функцию, входят в состав нуклеотидов Нуклеоти́ды— фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Мономерные единицы из которых состоит ДНК и РНК. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах. Эту картинку объясни 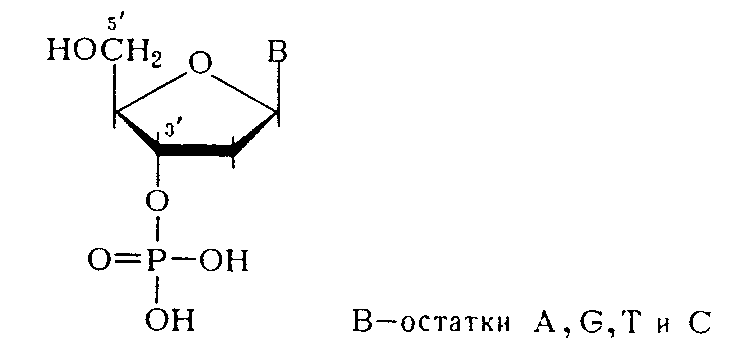 Нуклеиновые кислоты— высокомолекулярные соединения со строго определенной линейной последовательностью мононуклеотидов, носители генетической информации обо всех белках. 2 типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). пуриновые - а (А), (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U). Пентозы либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Пентозу соединяет с основанием N-гликозидная связь, Первичная структура ДНК- порядок чередования дезоксирибонуклеоЗИДмонофосфатов (дНМФ) в полинукпеотидной цепи. Вторичная структура- Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны. Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются за счёт водородных связей между А и Т (две связи) и между G и С (три связи).Правило Чаргаффа:« число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С)». Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль. Первичная структура РНК- порядок чередования рибонуклеоЗИДмонофосфатов (НМФ) в полинуклеотидной цепи. В РНК, как и в ДНК, нук-леотиды связаны между собой 3',5'-фосфодиэфирными связями Вторичная структура РНК- Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли - "шпильки", за счёт водородных связей между комплементарными азотистыми основаниями A-U и G-C. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например ионами Mg2+, В цитоплазме клеток присутствуют 3 типа рибонуклеиновых кислот - транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной масс, по функциональной активности. Отличия РНК и ДНК.. 2)Моносахарид (пентоза) в РНК представлен рибозой, в ДНК дезоксирибозой. 3)Азотистые основания в РНК- аденин, урацил, гуанин, цитозин; в ДНК- аденин, Тимин, гуанин, цитозин. 4)Первичная структура РНК нестабильна, в отличии от ДНК, т.к. имеет гидроксильную группу у 2`- углеводного атома рибозы. Переваривание нуклеопротеидов.Нуклеиновый компонент отделяется от белка в кислой среде желудка. Катаболизм нуклеиновых кислот начинается с гидролиза 3',5'-фосфодиэфирной связи под действием ферментов нуклеаз. 1. ДНКазы - расщепляют ДНК. 2. РНКазы - расщепляют РНК. Бывают эндонуклеазы (расщепляют внутренние 3'5'-фосфодиэфирные связи) и экзонуклеазы (отщепляют концевые мононуклеотиды). Встречается 2 типа экзонуклеаз: 3'-экзонуклеазы - отщепляют мононуклеотид с 3'-конца молекулы, и 5'-экзонуклеазы - отщепляют 5'-концевой мононуклеоти Пентозы, образующиеся в ходе катаболизма нуклеиновых кислот, могут быть утилизированы во II-м этапе ГМФ-пути. Азотистые основания также подвергаются дальнейшему катаболизму, но по-разному, в зависимости от их типа - пуриновых (аденина, гуанина) или пиримидиновых (тимина, цитозина и урацила).
продукт катаболизма пуриновых нуклеотидов - мочевая кислота. Её образование идёт путём гидролитического отщепления фосфатного остатка от нуклеотидов с помощью нуклеотидаз или фосфатаз 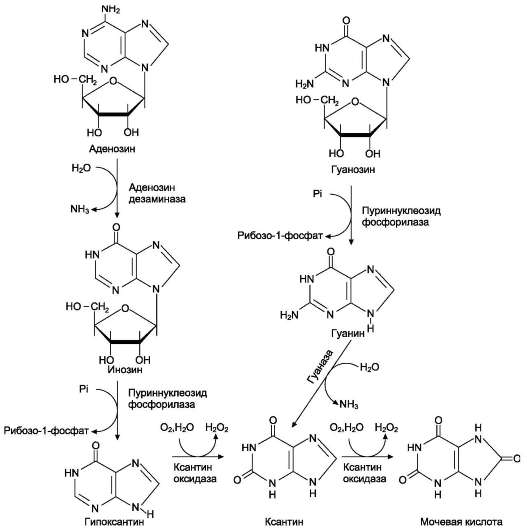 т 0,1 — 0,4 ммоль/л. Норма мочевой кислоты Повышение концентрации мочевой кислоты в крови называется гиперурикемией происходит отложение ее солей (уратов) в суставах и почках Это приводит к воспалительной реакции,боли. Такое заболевание называетсяподагра. Классическая подагра обусловлена тремя факторами — увеличенным синтезом мочевой кислоты, снижением содер¬жания в плазме уратсвязывающего белка и замедленным выведением с мочой. проявления подагры: 1 воспаления суствов); отложением кристаллов мононатриевой соли мочевой кислоты в суставе.. Для лечения подагры применяются: 1) ингибиторы фермента ксантиноксидазы. Например, аллопуринол - является конкурентным ингибитором фермента. Действие этого препарата приводит к повышению концентрации гипоксантина. Гипоксантин и его соли лучше растворимы в воде, и легче выводятся из организма.2) диетическое питание. Схему просто продиктовать словами 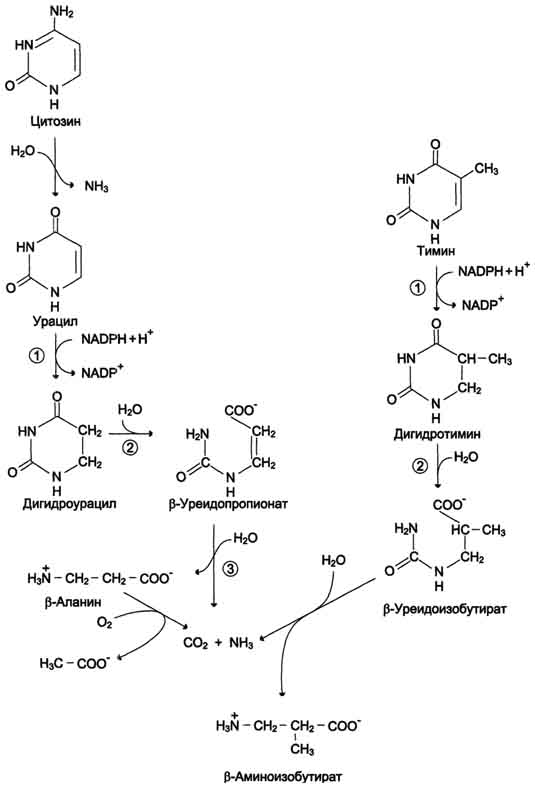 Катаболизм пиримидиновых оснований.1 дигидропиримидиндегидрогеназа; 2 - дигидропи-римидинциклогидролаза; 3 - уреидопропионаза. Часть β-аланина и β-аминоизобутирата тран-саминируется с α-кетоглутаратом и даёт малонил полуальдегид или метилмалонил полуальдегид соответственно, которые превращаются в ма-лонил-КоА и сукцинил-КоА и используются в соответствующих метаболических путях, либо окисляются до СО2 и Н2О. Частично β-амино-изобутират экскретируется с мочой.
Пуриновых синтез.
Источниками рибозо-5-фосфата могут быть: пентозофосфатный путь превращения глюкозы или катаболизм нуклеозидов Вторая реакция – это перенос NH2-группы глутамина на активированный атом С1 рибозо-5-фосфата с образованием 5'-фосфорибозиламина 5-фосфорибозиламин вовлекается в девять реакций, инозинмонофосфорная кислота (ИМФВ целом на синтез пуринового кольца затрачивается энергия 6 молекул АТФ. Превращение ИМФ в АМФ и ГМФ включает 2 стадии и идёт с затратой энергии Печень - основное место образования пури-новых нуклеотидов Схемой названия со всеми надн и ферментами 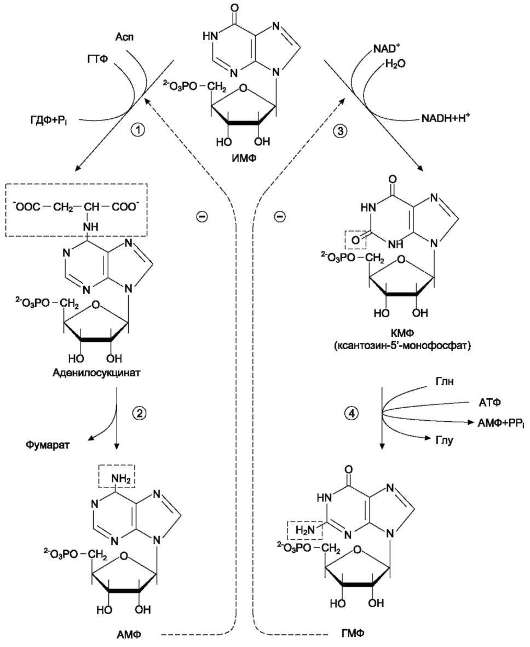 1 - аденилосукцинатсинтетаза; 2 - аденилосукциназа; 3 - ИМФ-дегидрогеназа; 4 - ГМФ-синтетаза. 1 - аденилосукцинатсинтетаза; 2 - аденилосукциназа; 3 - ИМФ-дегидрогеназа; 4 - ГМФ-синтетаза.аденилаткиназа катализирует реакцию: амф + атф → 2 адф, а гуанилаткиназа: гмф + атф → гдф + адф. гдф + атф → гтф + адф. ля синтеза пуринов такими ингибиторами являются АМФ и ГМФ. ГМФ блокирует первые две реакции синтеза ИМФ, а также ИМФ-дегидрогеназную реакцию. АМФ блокирует первую реакцию синтеза ИМФ и аденилосукцинатсинтетазную реакцию. Синтез пиримидиновых основанийпроисходит во всех клетках организма. В реакциях синтеза участвует аспарагиновая кислота, глутамин, СО2, затрачивается 2 молекулы АТФ.. Процесс протекает в цитозоле клеток. карбамоилфосфатсинтетазой II 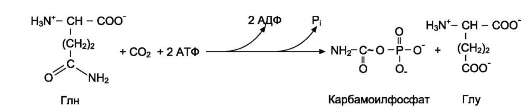 карбамо-илфосфат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт - дигидрооротат Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зависимой-дигидро-оротат-дегидрогеназой и превращается в свободное пиримидиновое основание - орото-вую кислоту, или оротат. В реакции с фосфорибозилдифосфатом (ФРДФ) к оротовой кислоте присоединяется рибозо-5-фосфат и образуется оротидилмонофосфат, при декарбоксилировании превращающийся в уридинмонофосфат (УМФ). Источником фосфорибозилдифосфата является первая из двух реакций синтеза фосфорибозиламина при образовании пуринов. 4. Синтез уридинтрифосфата.Синтез УТФ осуществляется из УМФ в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ. 5. Синтез цитидинтрифосфата.Образование цитидинтрифосфата (ЦТФ) происходит из УТФ с затратой энергии АТФ при участии глутамина, являющегося донором NH2-группы. НМФ-киназа катализирует следующую реакцию: умф + атф → удф + адф, а НДФ-киназа: удф + атф → утф + адф. ЦТФ синтетазакатализирует амидирование УТФ (рис. 10-14), осуществляя АТФ-зависимое замещение кетогруппы урацила на амидную группу глутамина с образованием цитидин-5 -трифосфата (ЦТФ). Регуляторным КАД-фермент. УМФ и УТФ аллостерически ингибируют, а ФРДФ активирует Дезоксирибонуклеозидтрифосфаты необходимы клетке для синтеза ДНК. 1. Реакция дефосфорилирования. В самом начале процесса происходит потеря рибонуклеозидтрифосфатами одной фосфатной группы и образуются АДФ, ГДФ, ЦДФ, УДФ. 2. Реакция восстановления. Во второй реакции фермент рибонуклеозид-редуктаза восстанавливает АДФ, ГДФ, ЦДФ, УДФ до дезоксирибонуклеоЗИДдифосфатов 3. Реакция фосфорилирования.После образования dАДФ, dГДФ, dЦДФ фосфорилируются, а dУДФ используется для синтеза тимидилового нуклеотида. Три дезоксирибонуклеотида– dАТФ, dГТФ, dЦТФ сразу после синтеза используются для синтеза ДНК. Однако известно, что в составе ДНК нет уридиловых нуклеотидов, поэтому dУДФ не превращается в dУТФ, а идет на образование тимидилового нуклеотида. Участие в этом принимает фермент тимидилатсинтаза.
(репликация, удвоение) ДНК происходит в S-фазу клеточного цикла, когда клетка готовится к делению Репликация проходит в три этапа:
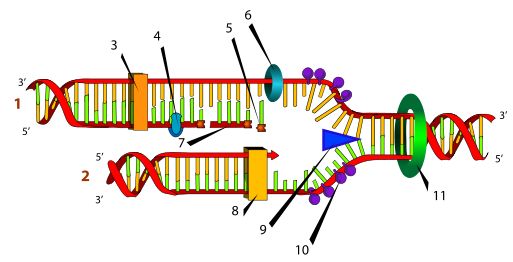
1.ДНК-топоизомеразы, находясь перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее расплетания и раскручивания. 2. ДНК-хеликазы, следуя за топоизомеразами, раскручивают и расплетают молекулу ДНК. 3. ДНК-связывающие белки (ДСБ) связывают расплетенные нити ДНК и стабилизируют их, не допуская обратного "слипания" друг с другом. 4. ДНК-полимераза δ (греч.: δ – дельта), согласовано со скоростью движения репликативной вилки, осуществляет синтез ведущей цепи дочерней ДНК в направлении 5'→ 5. Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (α- альфа ) и в направлении 5'→3' синтезирует праймер (РНК-затравку) . После этого фермент удаляется с нити ДНК.Вместо ДНК-полимеразы α к 3'-концу праймера присоединяется ДНК-полимераза ε. 6. ДНК-полимераза ε (греч.: ε – эпсилон) продолжает удлинять праймер, но в качестве субстрата встраивает дезоксирибонуклеотиды В результате образуется цельная нить из двух частей – РНК (т.е. праймер) и ДНК. ДНК-полимераза ε работает до тех пор, пока не встретит праймер предыдущего фрагмента Оказаки (синтезированный чуть ранее). После этого данный фермент удаляется с цепи. 7. ДНК-полимераза β (греч.: β – бета) встает вместо ДНК-полимеразы ε, движется в том же направлении (5'→3') и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, т.е. пока на его пути не встанет дезоксирибонуклеотид Связать результат свой работы и впереди стоящую ДНК фермент не в состоянии, поэтому он сходит с цепи. В результате на матрице материнской нити "лежит" фрагмент дочерней ДНК. Он называется фрагмент Оказаки. 8. ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки, т.е. 5'-конца отрезка, синтезированного ДНК-полимеразой ε, и 3'-конца цепи, встроенного ДНК-полимеразой β. |