1. Первичная структура белков
 Скачать 1.7 Mb. Скачать 1.7 Mb.
|

Белки плазмы кровиБелки являются важной составной частью крови и выполняют следующие функции: 1) определяют онкотическое давление; 2) обеспечивают вязкость крови; 3) обеспечивают свертываемость крови; 4) участвуют в регуляции кислотно-основного равновесия; 5) выполняют транспортную функцию (переносят липиды, НЭЖК, металлы, билирубин, гемоглобин, гор моны, лекарственные вещества); 6) обеспечивают иммунитет (антитела, интерферон и др.); 7) питательная функция (белки являются резервом аминокислот). Белки плазмы обычно делят на альбумины, глобулины и фибриноген. АльбуминыЭто простые, высокогидро фильные белки. Образуются в гепа тоцитах печени. Выполняют следующие функции:
Глобулиныразделены электрофорети чески на подгруппы. и -гло булины вырабатываются в ретикулоэндотели альной системе, в том числе купферовскими клетками печени. -глобулины состоят из глико- и липопротеидов. - глобулины участвуют в транспорте различных веществ. Они имеют самую высокую электрофоре-тическую подвиж ность. -глобулины состоят из глико-, липо- и металлопротеидов. Они выпол няют транспортную и другие функции. -глобулины с самой низкой электрофоретической подвижно стью. К этой группе относятся большинство защитных веществ крови, многие из которых обладают фер ментативной ак тив ностью. -глобулины синтезируются плазматическими клетками. Белки - ферменты1. Собственные ферменты плазмы крови, которые участвуют в свертыва нии крови, растворении внутрисосуди стых сгустков и т.д. Эти ферменты синтезируются в печени. 2. Клеточные ферменты освобождаются из клеток крови и клеток других тканей в результате есте ственного рас пада (лизиса). при гепатите - активность аланина минотрансферазы, арги назы, аспартат-сердце Белки - переносчики. Трансферрин является -глобулином. Может взаимодействовать с Сu2+ и Zn2+ , но главным образом связывает и переносит Fe3+ в различные ткани. Гаптоглобин является 2-глобулином, выполняет следующие функции:
Церулоплазмин является 2-глобулином, выполняет следующие функции: - является переносчиком и регулятором концентрации ионов меди в ор ганизме, Белки острой фазыЭто группа белков плазмы, содержание которых увеличивается в ответ на повреждение ткани, воспале ние, опухо левый процесс. Эти белки синтезиру ются в печени и являются гликопротеинами. К белкам ост рой фазы отно сятся:
Гиперпротеинемия– увеличение общего содержания белков плазмы.рвата диарея, потеря воды организмом, а следовательно, и плазмой приводит к повышению концентрации белка в крови (относительная гиперпротеинемия). При ряде патологических состояний может наблюдатьсяабсолютная гиперпротеинемия, обусловленная увеличением уровня γ-глобулинов: например, гиперпротеинемия в результате инфекционного или токсического раздражения системы макрофагов; Гипопротеинемия, или уменьшение общего кол-ва белка в плазме крови, наблюдается гл образом при снижении уровня альбуминов.. Содержание общего белка снижается до 30–40 г/л. Гипопротеинемия наблюдается при поражении печеночных кл (острая атрофия печени, токсический гепатит и др.). Кроме того, гипопротеинемия может возникнуть при резко увеличенной проницаемости стенок капилляров, при белковой недостаточности (поражение пищеварительного тракта, карцинома и др.).
остаточный азот крови (сумма всех азотсодержащих веществ крови после удаления из неё белков = Небелковый азот крови). Нормальное содержание 14,3 – 28,6 ммоль/ 1) мочевина (примерно 50% 2) АКты (около 25%), 3) креатин и креатинин(7,5%; 4) полипептиды, нуклеотиды и азотистые основания (5%; 5)мочевая кислота (4%; 6) аммиак и индикан (0,5%; Индикан представляет собой калиевую или натриевую соль индоксилсерной кислоты, образующейся в печени при обезвреживании индола Ретенционная азотемия развивается в результате недостаточного выделения с мочой азотсодержащих продуктов Она в свою очередь может быть почечной и внепочечной. При почечной ретенционной азотемии концентрация остаточного азота в крови увеличивается вследствие ослабления очистительной функции почек. 90%мочевины вместо 50%Внепочечные в свою очередь подразделяются на надпочечные и подпочечные Продукционная азотемияразвивается при избыточном поступлении азотсодержащих продуктов в кровь, как следствие усиленного распада тканевых белков при обширных воспалениях, ранениях, ожогах, кахексии и др. Нередко наблюдаются азотемии смешанного типа. Уреми́я— острое или хроническое самоотравление организма, обусловленное почечной недостаточностью; накопление в крови главным образом токсических продуктов азотистого обмена (азотемия), нарушения кислотно-щелочного и осмотического равновесия. Проявления: вялость, головная боль, рвота, диарея, кожный зуд, судороги, кома и др.
Гепатоцит имеет хорошо развитую систему эндоплазматического ретикулума ЭР как гладкую, так и шероховатую. Функции ЭР - синтез белков, (альбумины), или ферментов работающих в печени. синтезируются фосфолипиды, триглицериды и холестерол Функции печени:1.Пищеварительная–Она образует желчь, включающую воду (82%), желчные кислоты (12%), фосфатидилхолин (4%), холестерол (0,7%), прямой билирубин, белки.Желчь обеспечивает эмульгирование и переваривание жиров пищи, стимулирует перистальтику кишечника. 2. Экскреторнаяфункция, близка к пищеварительной – с помощью желчи выводятся билирубин, немного креатинина и мочевины, , холестерол.(в составе желчи) 3. Секреторная– печень синтез альбумина, белков свертывающей системы, липопротеинов, глюкозы, кетоновых тел, креатина. 4. Депонирующаядепо гликогена, мин. в-в, особенно железо, витамины A, D, K, B12 и фолиевая кислота. 5.Метаболическаяфункция – поддержание метаболического гомеостаза *Углеводный обмен.. Благодаря синтезу и распаду гликогена печень поддерживает конц-ию глюкозы в крови. Гликогена в печени30-100гр.при длительном голодании источником глюкозы яв-ся глюконеогенез из АК и глицерин. превращение гексоз (фруктозы, галактозы) в глюкозу. р-ии ПФ пути обеспечивают синтез НАДФН, необх-го для синтеза ж к-т и холестерола из глюкозы. *Липидный обмен. Если поступает избыток глюкозы, который не используется для синтеза гликогена и других синтезов, то она превращается в липиды – холестерол и триацилглицеролы. их удаление происходит при помощи ЛПОНП.. При сильном голодании синтезируются кетоновые тела которые яв-ся альтер. ист. Е *Белковый обмен. За 7 суток обновляются белки печени– альбумины, многие глобулины, ферменты крови, фибриноген и факторы свертывания крови. АК подвергаются реакциям с трансаминированием и дезаминированием, декарбоксилированию с образованием биогенных аминов. идет утилизация избыточного N и включение его в состав мочевины. *Пигментный обмен. Участие превращении гидрофобного билирубина в гидрофильную форму и секреция его в желчь 6. Обезвреживающаяфункция - биотрансформации подвергаются: а) стероидные и тиреоидные гормоны, инсулин, адреналин, б) продукты распада гемопротеинов (билирубин), в) продукты жизнед-ти микрофлоры, всасывающихся из толстого кишечника – кадаверин (производное лизина), путресцин (производное аргинина), крезол и фенол (производное фенилаланина и тирозина) и других токсинов, г)ксенобиотики (токсины, лекарственные вещества и их метаболиты).
путем аминирования или переаминирования пировиноградная кислота, являющаяся продуктом распада углеводов, может превратиться в аминокислоту -аланин. Кроме того, пировиноградная кислота в результате дальнейших превращений дает щавелевоуксусную (СООН—СН2—СО—СООН) и a-кетоглютаровую (СООН—СН2—СН2—СО—СООН) кислоты, из которых путем реакции аминирования и переаминирования соответственно образуются аспарагиновая и глютаминовая аминокислоты. углеводы в животном организме могут синтезироваться из продуктов окисления белков. Углеводы образуются из тех аминокислот, которые при своем дезаминировании превращаются в кетокислоты. СВЯЗЬ МЕЖДУ ОБМЕНОМ УГЛЕВОДОВ И ЖИРОВ Единство в обмене углеводов и жиров доказывается возникновением общих промежуточных продуктов распада. При распаде углеводов образуется пировиноградная кислота, а из нее -активная уксусная кислота -ацетил-КоА, который может быть использован в синтезе жирных кислот. Последние при своем распаде дают ацетил-КоА. Для синтеза нейтрального жира необходим кроме жирных кислот и глицерин. Глицерин также может синтезироваться из продуктов распада углеводов, а именно, из фосфоглицеринового альдегида и фосфодиоксиацетона. И наоборот, при распаде глицерина могут образовываться фосфотриозы. СВЯЗЬ МЕЖДУ ОБМЕНОМ БЕЛКОВ И ЖИРОВ Многие заменимые аминокислоты могут синтезироваться из промежуточных продуктов расщепления жиров. Возникающий при распаде жирных кислот цетил-КоА вступает в конденсацию с щавелевоуксусной кислотой и через цикл трикарбоновых кислот приводит к образованию a-кетоглютаровой кислоты. Кетоглютаровая кислота в результате аминирования или переаминирования переходит в глютаминовую. Глицерин, входящий в состав нейтральногo жира, окисляется в глицериновую кислоту и в дальнейшем превращается в пировиноградную, а последняя используется для синтеза заменимых аминокислот.Использование белков для синтеза жира осуществляется через образование ацетил-КоА. Далее ацетил-КоА может быть использован для синтеза жирных кислот. Глицерин образуется лишь за счет тех аминокислот, которые способны превращаться в пировиноградную кислоту. 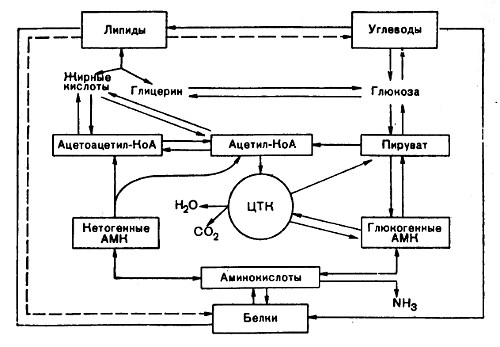
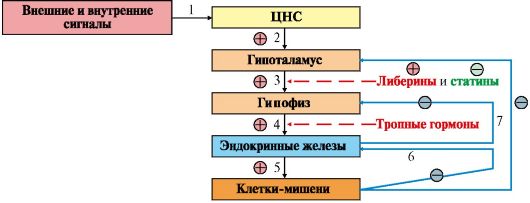
Интеграция – это объединение элементов системы в единое целое. Координация (соподчинение) – это подчинение менее важных элементов системы более важным элементам. Интеграция и координация – это две стороны процесса регуляции. Различают:
Механизмы клеточной ауторегуляции 1. Компартментализация (мембранный механизм). Роль мембран состоит в следующем: а) мембраны делят клетки на отсеки и в каждом из них осуществляются свои процессы; б) мембраны обеспечивают активный транспорт и регулируют потоки молекул в клетке и из клетки; в) в мембраны встроены ферменты; г) мембраны защищают клетку от внешних воздействий. Воздействием на функции мембран клетка может регулировать тот или иной процесс. 2. Изменение активности ферментов. 3. Изменение количества ферм Классификация межклеточных регуляторов Анатомо-физиологическая: а) Гормоны – межклеточные регуляторы, доставляемые к клеткам-мишеням током крови. Вырабатываются в эндокринных железах б) Нейрогормоны вырабатываются нервными клетками и выделяются в синаптическую щель. Нейрогормоны делятся на медиаторы и модуляторы. Медиаторы обладают непосредственным пусковым эффектом. Модуляторы изменяют эффект медиаторов. Примерами медиаторов являются ацетилхолин и норадреналин; модуляторов – -ааминомасляная кислота, дофамин. в) Локальные гормоны – это межклеточные регуляторы, действующие на близлежащие к месту их синтеза клетки. Пример: гормоны, производные жирных кислот. Классификация по широте действия: а) Гормоны универсального действия действуют на все ткани организма (например, катехоламины, глюкокортикостероиды). б) Гормоны направленного действия действуют на определенные органы-мишени (например, АКТГ действует на кору надпочечников). Классификация по химическому строению: а)Белково-пептидные гормоны
б)Производные аминокислот:
в)Липидные гормоны:
Табличку продиктуй! На стимулирующие или тормозящие стимулы из ЦНС секретируются стимулирующие или ингибирующие рилизинг-факторы, которые носят название либерины или статины соответственно. Эти нейрогормоны с кровотоком достигают аденогипофиза, где стимулируют (либерины) или ингибируют (статины) биосинтез и секрецию тропных гормонов. Тропные гормоны воздействуют на периферические железы, стимулируя выделение соответствующих периферических гормонов
Рецепторы – это белковые молекулы, специфически связывающие данный гормон, в результате чего возникает какой-либо эффект. Гормон начинает свое действие с соединения с рецептором, образуя гормон-рецепторный комплекс. Рецепторы могут находиться внутри клетки, а также на клеточной мембране. Механизм действия гормонов через внутриклеточные рецепторы. Гормон проникает в клетку, связывается с рецептором. Образованный таким образом гормон-рецепторный комплекс перемещается в ядро и действует на генетический аппарат клетки. В результате меняется процесс транскрипции, а в дальнейшем, синтез белков. Таким образом, данные гормоны влияют на количество ферментов в клетке. Механизм действия гормонов через рецепторы плазматических мембран В этом случае гормон не проникает в клетку, а взаимодействует с рецептором на поверхности мембраны.
Хорошо изученными вторыми посредниками являются циклические нуклеотиды (цАМФ, цГМФ) и Ca2+. Механизм действия гормонов через цАМФ Когда соответствующий гормон связывается с рецептором, в мембране активируется фермент аденилатциклаза, который из АТФ образует цАМФ. цАМФ является аллостерическим активатором протеинкиназы, которая фосфорилирует белки и изменяет их свойства. Содержание цАМФ в клетке увеличивают: глюкагон, катехоламины (через -рецепторы), антидиуретический гормон, гистамин (Н2-рецепторы), простагландин-Е, простациклин, тиреотропный гормон, АКТГ, холерный токсин. Содержание цАМФ в клетке снижают: ацетилхолин (М-холинорецепторы), катехоламины (2-рецепторы), соматостатин, ангиотензин-II, опиаты, коклюшный токсин. Механизм действия гормонов через Са2+ Когда гормон связывается с рецептором, в мембране открывается кальциевый канал. В результате содержание кальция в клетке возрастает. Кальций связывается с белком клеток – кальмодулином, образуется комплекс, который может действовать непосредственно на белки, вызывая эффекты, или действовать на кальмодулин-зависимую протеинкиназу. Эта протеинкиназа фосфорилирует белки, в результате изменяются их свойства. Са2+ в качестве второго посредникавыполняет те же функции, что и цАМФ, за исключением того, что в гладких мышцах вызывает сокращение, тромбоцитах – агрегацию. Содержание кальция в клетке повышают: катехоламины через 1-рецепторы, ацетилхолин через М-холинорецепторы, гистамин через Н1-рецепторы, тромбоксан, ангиотензин-II.
Инсулин является белкого-пептидным гормоном с молекулярной массой 5700. Синтезируется в В-клетках поджелудочной железы из проинсулина. проинсулин, который транспортируется в комплекс Гольджи, далее в цистернах которого происходит так называемое созревание инсулина. В процессе созревания из молекулы проинсулина с помощью специфических эндопептидаз вырезается C-пептид Скорость секреции инсулина зависит от концентрации глюкозы в крови: при повышении концентрации секреция инсулина увеличивается, а при снижении – уменьшается. Молекула инсулина образована двумя полипептидными цепями, содержащими 51аминокислотный остаток: A-цепь состоит из 21 аминокислотного остатка, B-цепь образована 30 аминокислотными остатками. Полипептидные цепи соединяются двумя дисульфидными мостиками через остатки цистеина, третья дисульфидная связь расположена в A-цепи. 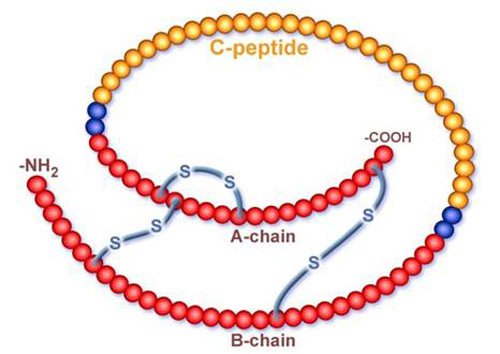 Секрецию инсулина также усиливают: глюкагон, секретин, холецистокинин, СТГ и пища, богатая белками. Рецепторы для инсулина находятся на клеточной мембране, Главными мишенями для инсулина являются мышцы, печень, жировая ткань, фибробласты и лимфоциты. Головной мозг не зависит от инсулина.
усиливает поглощение клетками аминокислот (особенно лейцина и валина);
Может быть 2 причины сахарного диабета:
Различают инсулинзависимый(юношеский, ювенильный) и инсулиннезависимый (стабильный) сахарный диабет. При инсулинзависимом диабете наблюдается абсолютная недостаточность инсулина, и жизнь больных зависит от инъекции инсулина. При инсулиннезависимом диабете наблюдается относительная недостаточность инсулина, поддержание глюкозы на нормальном уровне достигается сахаропонижающими средствами, инъекции инсулина не требуются. |