Патфиз ч.2. Патфиз ч. Гл. 11 Нарушения водного обмена Гл. 12 Нарушения ионного обмена
 Скачать 7.84 Mb. Скачать 7.84 Mb.
|

|
В организме человека постоянно происходят мутации. Их суммарное количество в расчёте на один клеточный цикл составляет примерно 1 СИСТЕМА ИММУНОБИОЛОГИЧЕСКОГО НАДЗОРА Биологическое значение системы иммунобиологического надзора ИБН заключается в контроле (надзоре) за индивидуальным и однородным клеточномолекулярным составом организма. Обнаружение носителя чужеродной генетической или антигенной информации (молекулы, вирусы, клетки или их фрагменты) сопровождается его инактивацией, деструкцией и, как правило, элиминацией. При этом клетки иммунной системы способны сохранять «память» о данном агенте. Повторный контакт такого агента с клетками системы ИБН вызывает развитие эффективного ответа, который формируется при участии как специфических — иммунных механизмов защиты, так и неспецифических факторов резистентности организма (рис. 16–1). 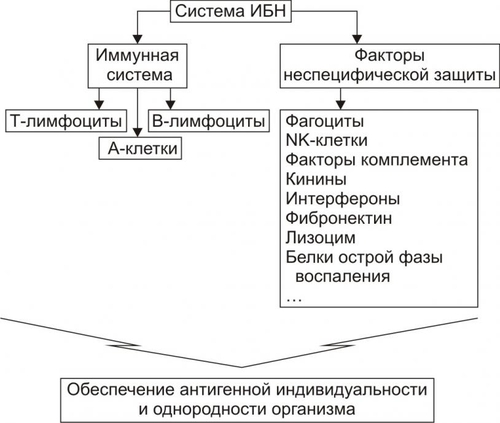 Рис. 16–1. Структура системы иммунобиологического надзора организма. NK — natural killers (естественные киллеры). А-клетки — антигенпредставляющие клетки. К числу основных в системе представлений о механизмах надзора за индивидуальным и однородным антигенном составе организма относят понятия об Аг, иммунитете, иммунной системе и системе факторов неспецифической защиты организма. АНТИГЕНЫ Инициальным звеном процесса формирования иммунного ответа является распознавание чужеродного агента — антигена (Аг). Происхождение этого термина связано с периодом поиска агентов, веществ или «тел», обезвреживающих факторы, вызывающие болезнь, а конкретно речь шла о токсине дифтерийной палочки. Эти вещества назвали вначале «антитоксинами», а вскоре был введён более общий термин «антитело». Фактор же, приводящий к образованию «антитела» обозначили как «антиген». Антиген — вещество экзо- или эндогенного происхождения, вызывающее развитие иммунных реакций (гуморального и клеточного иммунных ответов, реакций гиперчувствительности замедленного типа и формирование иммунологической памяти). Учитывая способность Аг вызывать толерантность, иммунный или аллергический ответ их называют ещё, соответственно, толерогенами, иммуногенами или аллергенами соответственно. Различный результат взаимодействия Аг и организма (иммунитет, аллергия, толерантность) зависит от ряда факторов: от свойств самого Аг, условий его взаимодействия с иммунной системой, состояния реактивности организма и других (рис. 16–2). 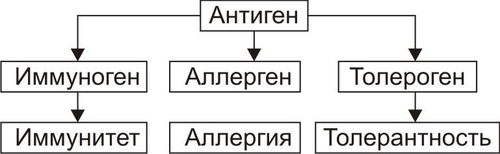 Рис. 16–2. Потенциальные эффекты антигена в организме. АНТИГЕННАЯ ДЕТЕРМИНАНТА Образование АТ и сенсибилизацию лимфоцитов вызывает не вся молекула Аг, а только особая его часть — антигенная детерминанта, или эпитоп. У большинства белковых Аг такую детерминанту образует последовательность из 4–8 аминокислотных остатков, а у полисахаридных Аг — 3–6 гексозных остатков. Число же детерминант у одного Аг может быть различным. Так, у яичного альбумина их не менее 5, у дифтерийного токсина — минимум 80, у тиреоглобулина — более 40. ВИДЫ АНТИГЕНОВ В соответствии со структурой и происхождением Аг подразделяют на несколько видов. • В зависимости от структуры различают белковые и небелковые Аг. † Белки или сложные вещества (гликопротеины, нуклеопротеины, ЛП). Их молекулы могут иметь несколько различных антигенных детерминант; † Вещества, не содержащие белка, называют гаптенами. К ним относятся многие моно, олиго и полисахариды, липиды, гликолипиды, искусственные полимеры, неорганические вещества (соединения йода, брома, висмута), некоторые ЛС. Сами по себе гаптены неиммуногенны. Однако после их присоединения (как правило, ковалентного) к носителю —молекуле белка или белковым лигандам клеточных мембран они приобретают способность вызывать иммунный ответ. Молекула гаптена обычно содержит лишь одну антигенную детерминанту. • В зависимости от происхождения различают экзогенные и эндогенные Аг. † Экзогенные Аг подразделяют на инфекционные и неинфекционные. ‡ Инфекционные и паразитарные Аг (вирусов, риккетсий, бактерий, грибов, одно и многоклеточных паразитов). ‡ Неинфекционные (чужеродные белки; белоксодержащие соединения; Аг и гаптены в составе пыли, пищевых продуктов, пыльцы растений, ряда ЛС). † Эндогенные Аг (аутоантигены) появляются при повреждении белков и содержащих белок молекул собственных клеток, неклеточных структур и жидкостей организма, при конъюгации с ними гаптенов, в результате мутаций, приводящих к синтезу аномальных белков, при сбоях иммунной системы. Другими словами, во всех случаях когда Аг распознаётся как чужеродный. ИММУНИТЕТ В иммунологии термин «иммунитет» применяют в трёх значениях. • Для обозначения состояния невосприимчивости организма к воздействию носителя чужеродной генетической или антигенной информации (бактерии, вирусы, риккетсии, паразиты, грибы, клетки чужеродного трансплантата, опухолей и др.). • Для обозначения реакций системы ИБН против Аг. • Для обозначения физиологической формы иммуногенной реактивности организма, наблюдающейся при контакте клеток иммунной системы с генетически или антигенно чужеродной структурой. В результате эта структура подвергается деструкции и, как правило, элиминируется из организма. ИММУННАЯ СИСТЕМА Иммунная система — комплекс органов и тканей, содержащих иммунокомпетентные клетки и обеспечивающая антигенную индивидуальность и однородность организма путём обнаружения и, как правило, деструкции и элиминации из него чужеродного Аг. Иммунная система состоит из центральных и периферических органов. • К центральным (первичным) органам относят костный мозг и вилочковую железу. В них происходит антигеннезависимое деление и созревание лимфоцитов, которые впоследствии мигрируют в периферические органы иммунной системы. • К периферическим (вторичным) органам относят селезёнку, лимфатические узлы, миндалины, лимфоидные элементы ряда слизистых оболочек. В этих органах происходят как антигеннезависимая, так и антигензависимая пролиферация и дифференцировка лимфоцитов. Как правило, зрелые лимфоциты впервые контактируют с Аг именно в периферических лимфоидных органах. † Заселение периферических органов иммунной системы T и B-лимфоцитами, поступающими из центральных органов иммунной системы, происходит не хаотически. Каждая популяция лимфоцитов мигрирует из кровеносных сосудов в определённые лимфоидные органы и даже в различные их регионы. Так, B-лимфоциты преобладают в селезёнке (в её красной пульпе, а также по периферии белой) и пейеровой бляшке кишечника (в центрах фолликулов), а T-лимфоциты — в лимфатических узлах (в глубоких слоях их коркового вещества и в перифолликулярном пространстве). † В организме здорового человека в процессе лимфопоэза образуется более 109 разновидностей однородных клонов лимфоцитов. При этом каждый клон экспрессирует только один вид специфического антигенсвязывающего рецептора. Большинство лимфоцитов периферических органов иммунной системы не закрепляются в них навсегда. Они постоянно циркулируют с кровью и лимфой как между различными лимфоидными органами, так и во всех других органах и тканях организма. Такие лимфоциты получили название рециркулирующих. † Биологический смысл рециркуляции T и B-лимфоцитов: ‡ Вопервых, осуществление постоянного надзора за антигенными структурами организма. ‡ Вовторых, реализация межклеточных взаимодействий (кооперация) лимфоцитов и мононуклеарных фагоцитов, что необходимо для развития и регуляции иммунных реакций. ИММУНОКОМПЕТЕНТНЫЕ КЛЕТКИ К иммунокомпетентным клеткам относятся T- и B-лимфоциты, NK-клетки и антигенпредставляющие клетки (рис. 16–3). 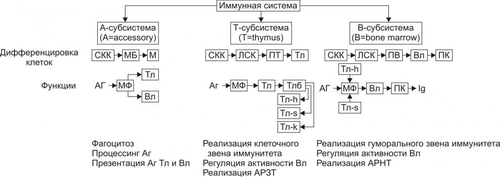 Рис. 16–3. Организация и функции системы иммунокомпетентных клеток. СКК — стволовая кроветворная клетка, МБ —миелобласт, М —миелоцит, ЛСК — лимфопоэтическая клетка, ПТ — преT-лимфоцит, Тл — T-лимфоцит, Вл — B-лимфоцит, ПК — плазматическая клетка, Мф — макрофаг, Тлб — T-лимфоцит в состоянии бласттранcформации, Тлh — T-лимфоцитхелпер, Тлs — T-лимфоцитсупрессор, Тлk — T-лимфоциткиллер, АРЗТ — аллергическая реакция замедленного типа, АРНТ — аллергическая реакция немедленного типа. Т-лимфоциты развиваются в тимусе из клеток-предшественниц. В-лимфоциты дифференцируются в печени плода и костном мозге взрослого организма. NK-клетки образуются из предшественников лимфоидных клеток в костном мозге. Лимфоциты, как и другие лейкоциты, на своей поверхности экспрессируют большое количество различных молекул, по которым при помощи моноклональных АТ идентифицируют их принадлежность к конкретной клеточной популяции. Чаще всего с этой целью выявляют дифференцировочные антигены (CD), являющиеся специфичными клеточными маркёрами. Идентификация клеточных маркёров при помощи АТ используется в проточной цитометрии для сортировки и подсчёта количества клеток в исследуемых популяциях. B-ЛИМФОЦИТЫ Эта субсистема образована различными клонами B-лимфоцитов. Название субсистемы отражает то обстоятельство, что лимфоциты, представляющие её, формируются у птиц в сумке (bursa) Фабрициуса (впервые B-лимфоциты были выявлены в лимфоидных органах птиц). У человека подобной бурсы нет, B-лимфоциты созревают в костном мозге, а также, возможно, в пейеровых бляшках, миндалинах, определённых зонах селезёнки и лимфоузлов. B-лимфоциты берут начало от стволовых кроветворных клеток костного мозга. B-лимфоциты обеспечивают реализацию эффекторного звена гуморального иммунного ответа. В мембране B-лимфоцита присутствует рецептор Аг — мономер IgM. Из красного костного мозга B-лимфоциты мигрируют в тимус-независимые зоны лимфоидных органов. Продолжительность жизни большинства B-лимфоцитов не превышает десяти дней, если они не активируются Аг. Зрелые В-лимфоциты (плазматические клетки) вырабатывают АТ — Ig всех известных классов. CD19, CD20 и CD22 — основные маркёры, используемые для идентификации B-клеток. В процессе формирования B-клеток выделяют антигеннезависимую и антигензависимую стадии. Антигеннезависимая стадия Антигеннезависимая стадия созревания B-лимфоцитов происходит под контролем локальных клеточных и гуморальных сигналов от микроокружения преB-лимфоцитов и не определяется контактом с Аг. На этой стадии происходит формирование отдельных пулов генов, кодирующих синтез Ig, а также экспрессия этих генов. Однако, на цитолемме преB-клеток ещё нет поверхностных рецепторов — Ig, компоненты последних находятся в цитоплазме. Образование B-лимфоцитов из преB-лимфоцитов сопровождается появлением на их поверхности первичных Ig, способных взаимодействовать с Аг. Только на этом этапе B-лимфоциты попадают в кровоток и заселяют периферические лимфоидные органы. Сформировавшиеся молодые B-клетки накапливаются в основном в селезёнке, а более зрелые — в лимфатических узлах. Антигензависимая стадия Антигензависимая стадия развития B-лимфоцитов начинается с момента контакта этих клеток с Аг (в том числе — аллергеном). В результате происходит активация B-лимфоцитов, протекающая в два этапа: пролиферации и дифференцировки. • Пролиферация B-лимфоцитов обеспечивает два важных процесса: † Увеличение числа клеток, дифференцирующихся в продуцирующие АТ (Ig) B-клетки (плазматические клетки). По мере созревания B-клеток и их превращения в плазматические клетки происходит интенсивное развитие белоксинтезирующего аппарата, комплекса Гольджи и исчезновение поверхностных первичных Ig. Вместо них продуцируются уже секретируемые (т.е. выделяемые в биологические жидкости — плазму крови, лимфу, СМЖ и др.) антигенспецифические АТ. Каждая плазматическая клетка способна секретировать большое количество Ig — несколько тысяч молекул в секунду. Процессы деления и специализации B-клетки осуществляются не только под влиянием Аг, но и при обязательном участии T-лимфоцитовхелперов, а также выделяемых ими и фагоцитами цитокинов — факторов роста и дифференцировки; † Образование В-лимфоцитов иммунологической памяти. Эти клоны B-клеток представляют собой долгоживущие рециркулирующие малые лимфоциты. Они не превращаются в плазматические клетки, но сохраняют иммунную «память» об Аг. Клетки памяти активируются при повторной их стимуляции тем же самым Аг. В этом случае B-лимфоциты памяти (при обязательном участии T-eeетокхелперов и ряда других факторов) обеспечивают быстрый синтез большого количества специфических АТ, взаимодействующих с чужеродным Аг, и развитие эффективного иммунного ответа или аллергической реакции. T-ЛИМФОЦИТЫ Субсистема T-лимфоцитов представлена различными клонами T-лимфоцитов. Их пролиферация и дифференцировка происходит под контролем вилочковой железы. В связи с этим их обозначают как T-клетки, или тимусзависимые лимфоциты. T-клетки, как и B-лимфоциты, развиваются из стволовых кроветворных клеток костного мозга. Отсюда в виде клеток–предшественниц T-лимфоциты попадают с кровью в тимус, где происходит их антигеннезависимое созревание, сопровождающееся экспрессией на цитолемме специфических (у каждого лимфоцита своего) рецепторов. Тлимфоциты ответственны за реализацию клеточного звена иммунного ответа, а также участвуют в регуляции гуморального иммунного ответа. Т-клетки состоят из функциональных подтипов CD4+ и CD8+. Тхелперы (TH) — CD4+ Т-клетки. При активации синтезируют и секретируют цитокины (ИЛ2, ИЛ4, ИЛ5, ИЛ6, Цитотоксические T-лимфоциты (TC) — CD8+ Т-клетки, уничтожают инфицированные вирусом, опухолевые и чужеродные клетки при помощи цитолитического белка — перфорина. Взаимодействуют с молекулой MHC класса I плазматической мембраны клетки–мишени. Tсупрессоры (TS) — представители CD8+ Т-клеток — регулируют интенсивность иммунного ответа, подавляя активность TH клеток; предотвращают развитие аутоагрессивных иммунных реакций; защищают организм от нежелательных последствий иммунной реакции, от чрезмерного воспаления и аутоагрессии. NK-КЛЕТКИ NK-клетки (МНСнерестригированные киллеры, естественные киллеры) составляют до 15% всех лимфоцитов крови. Они не имеют поверхностных детерминант, характерных для T- и B-лимфоцитов, не имеют рецептора Т-лимфоцитов. В типичных NK-клетках экспрессируются дифференцировочные Аг CD2, СD7, CD56 и CD16 (рецептор Fcфрагмента IgG). В плазматической мембране активированных NK-клеток появляется гликопротеин CD69. NK-клетки распознают и уничтожают опухолевые и вирус-инфицированные клетки. Механизм распознавания неясен. Существует представление о наличии поверхностноклеточных молекул, защищающих клетки организма от цитотоксического действия NK-eлеток. Примером служит продукт гена HLAC. Распознавание рецептором NK-eлетки этой молекулы тормозит цитотоксическую активность NK-eлеток и таким образом защищает клетку, экспрессирующую HLAC. Модификация продукта гена HLAC вирусами или связанными с опухолью молекулами приводит к уничтожению этой клетки NK-eлеткой. NK-eлетки, располагая рецептором IgG (CD16), способны также взаимодействовать с клетками, окружёнными молекулами IgG, и уничтожать их (феномен АТзависимой цитотоксичности). Активированные NK-клетки выделяют Цитолиз. В отличие от цитотоксических Tлимфоцитов, способность NK-клеток к цитолизу не связана с необходимостью распознавания молекул MHC на поверхности мишени. NK-клетки уничтожают клетку–мишень не путём фагоцитоза, а (после установления с ней прямого контакта) при помощи перфорина. Гуморальная регуляция. Активность NK-клеток регулируется цитокинами. Участие в антителозависимом клеточно-опосредованном цитолизе. NK-клетки, наряду с макрофагами, нейтрофилами и эозинофилами, участвуют также и в АТ-зависимом клеточно-опосредованном цитолизе. Для этого NK-клетки экспрессируют на своей поверхности рецептор Fcфрагмента IgG (CD16). Fcфрагмент этих АТ взаимодействует с рецептором Fcфрагмента, встроенным в плазматическую мембрану NK-клетки. АНТИГЕНПРЕДСТАВЛЯЮЩИЕ КЛЕТКИ Антигенпредставляющие клетки (А-субсистема на рис. 16-3) присутствуют преимущественно в коже, лимфатических узлах, селезёнке и тимусе. К ним относятся макрофаги, дендритные клетки, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерханса, Мклетки в лимфатических фолликулах пищеварительного тракта, эпителиальные клетки вилочковой железы. Эти клетки захватывают, перерабатывают и представляют Аг (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают ИЛ1 и другие цитокины, секретируют простагландин E2 (PGE2), угнетающий иммунный ответ. Фагоцитарную и цитолитическую активность макрофагов усиливает |