Основы биологической химии предисловие
 Скачать 7.85 Mb. Скачать 7.85 Mb.
|

Глава 4. НУКЛЕИНОВЫЕ КИСЛОТЫ
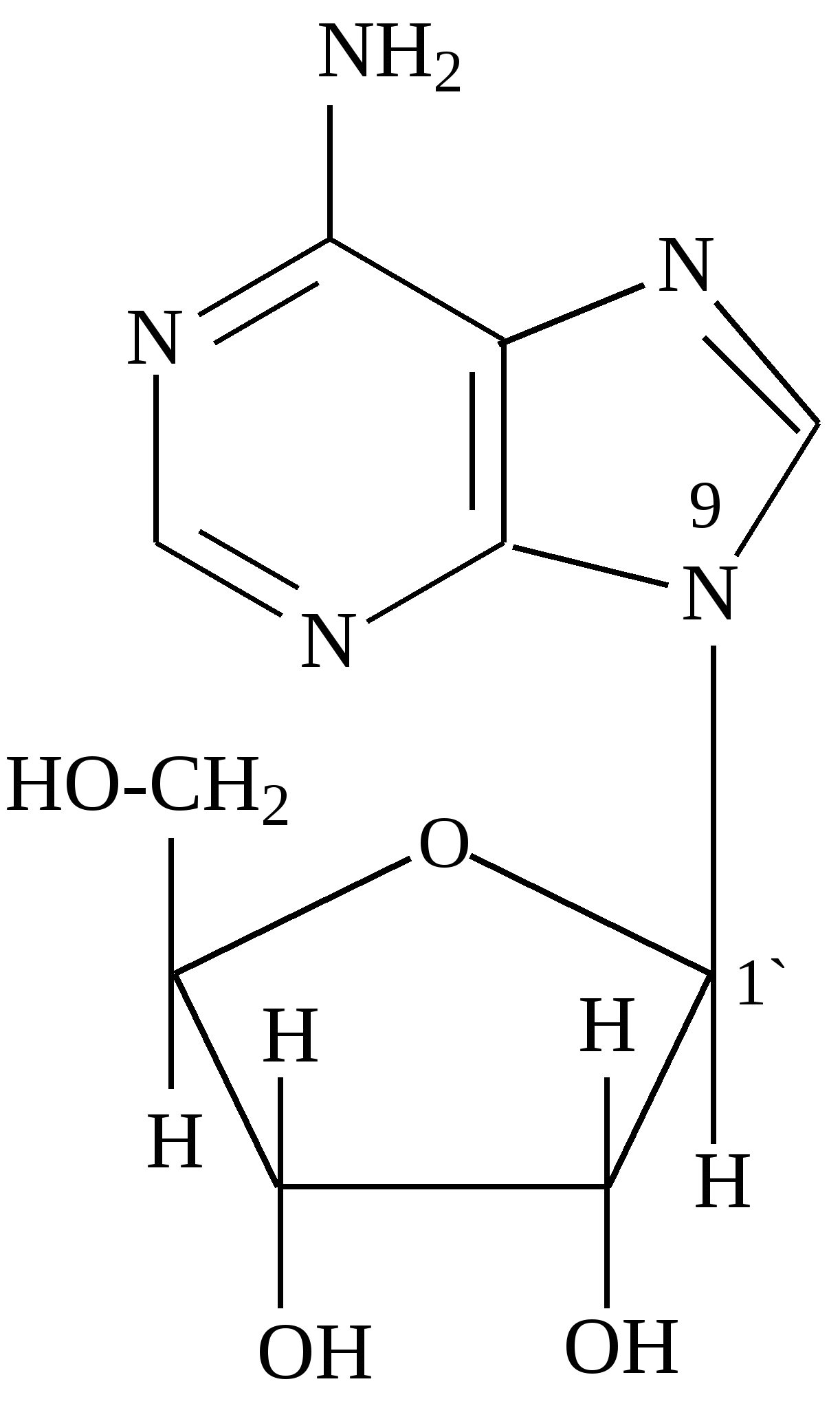
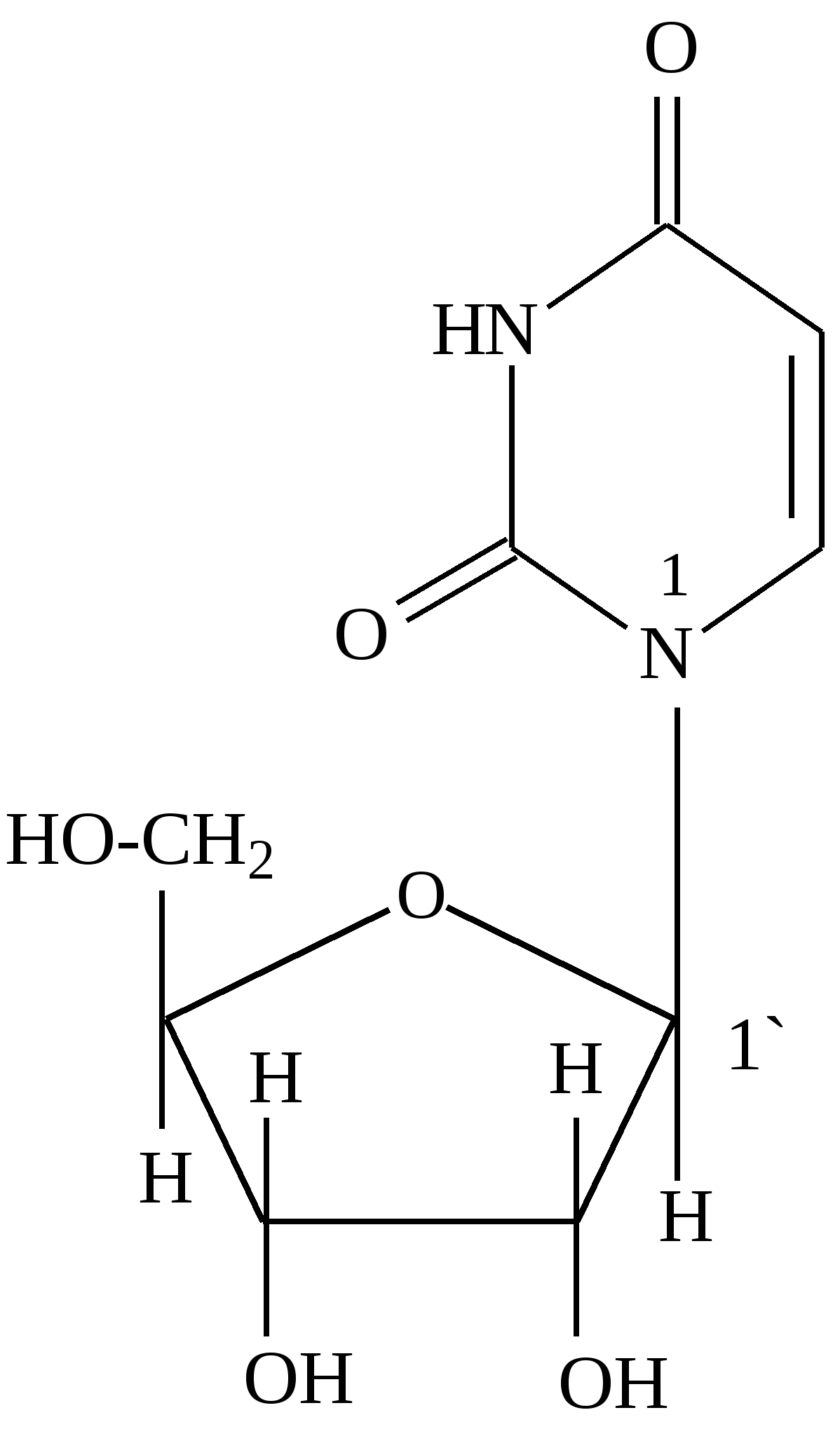
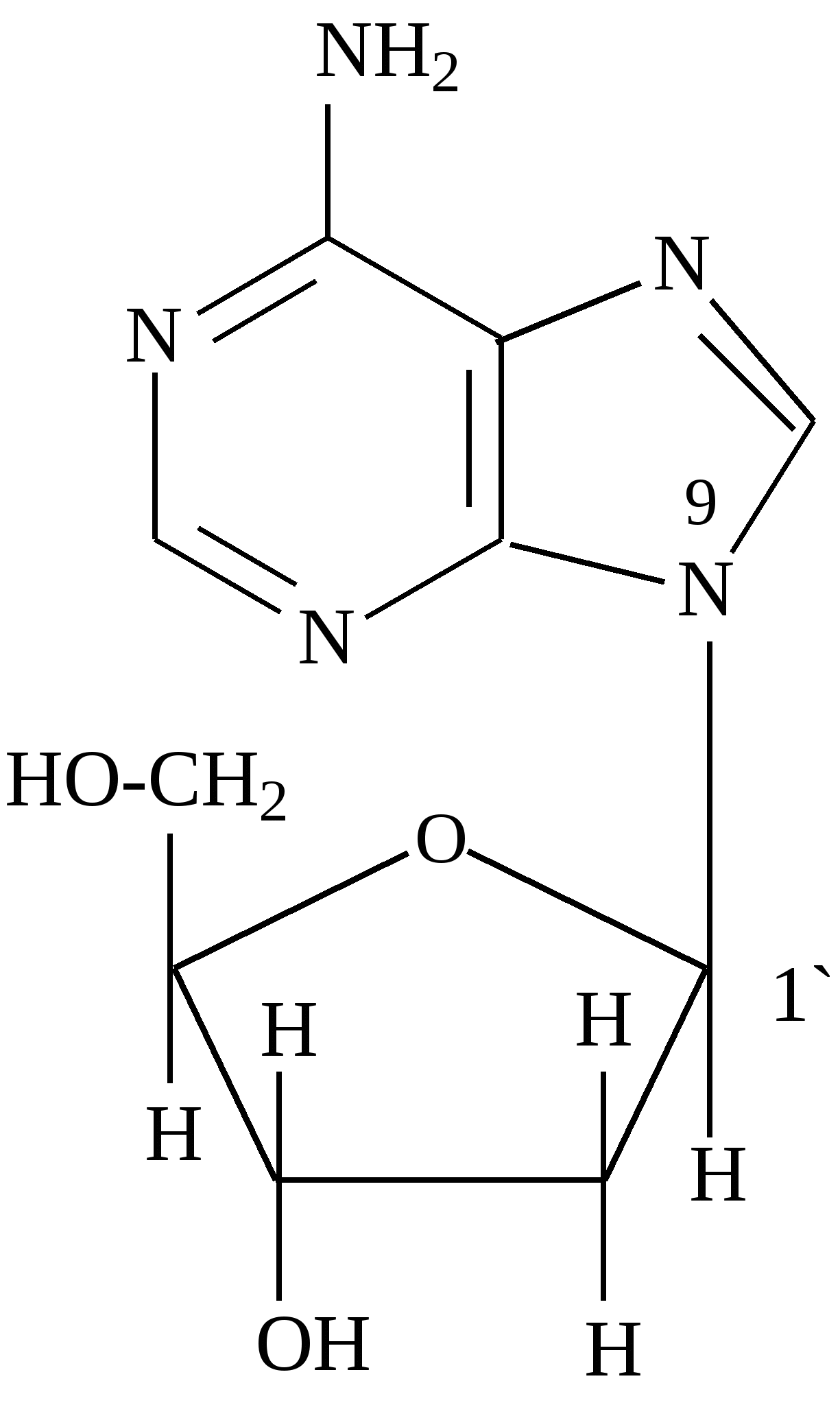
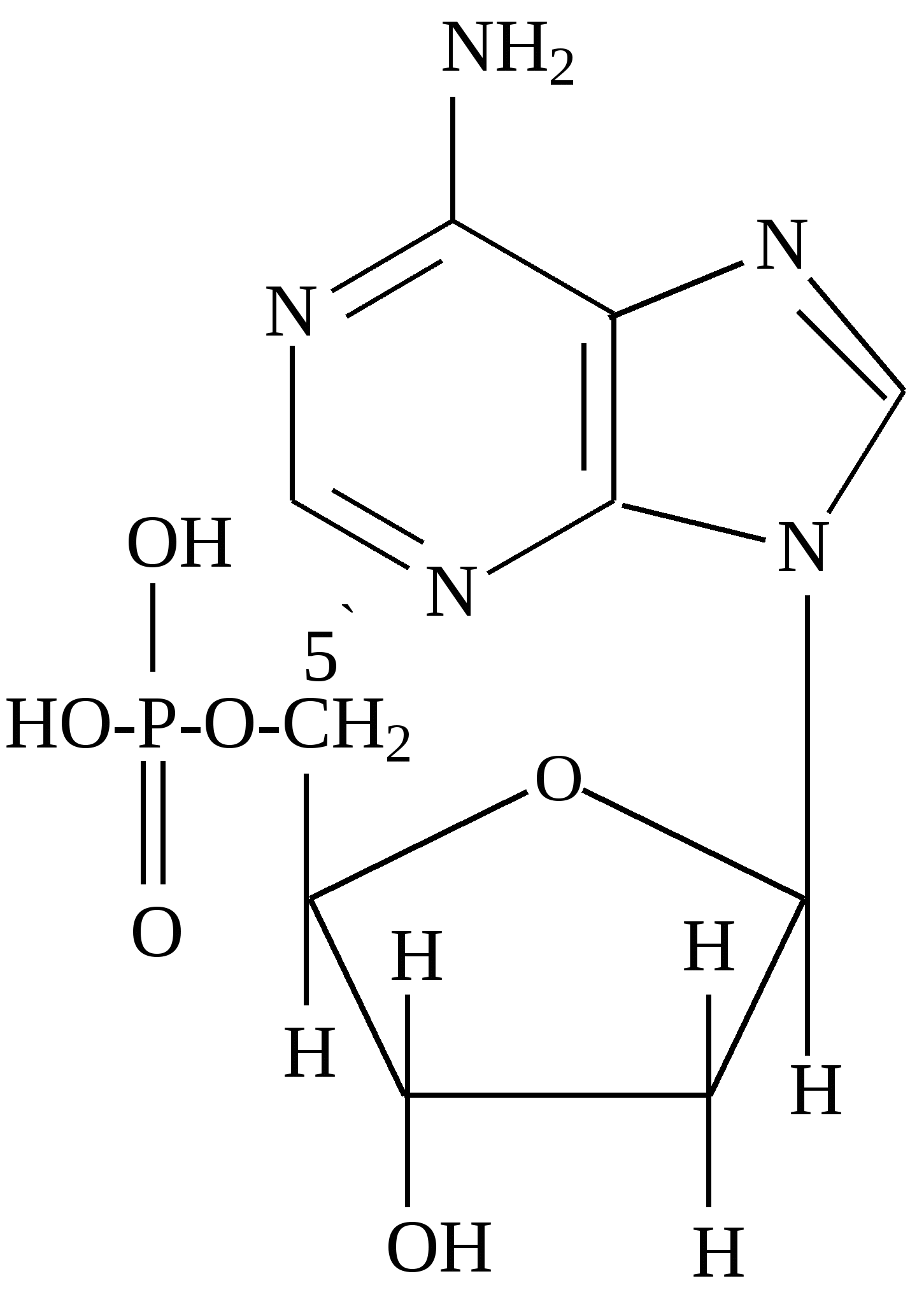
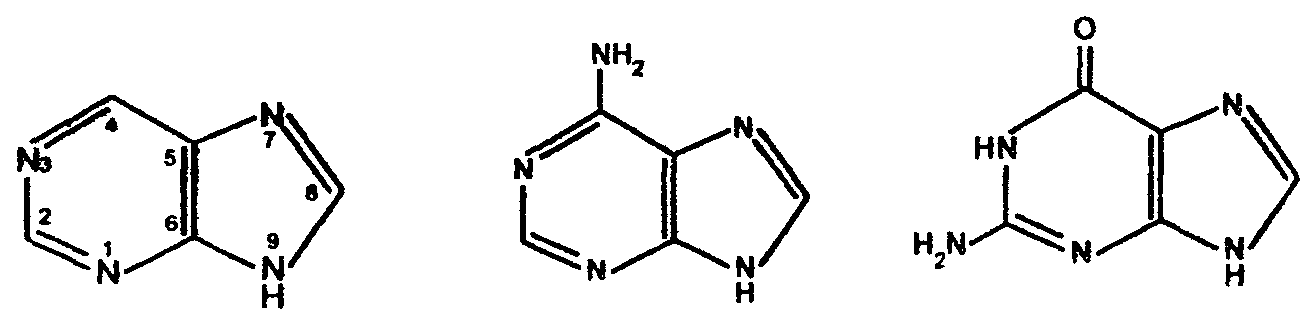
В фазе покоя клетки хроматин равномерно распределен по всему объему ядра и не обнаруживается обычными микроскопическими методами. В фазе деления клетки хроматин образует компактные частицы - хромосомы, которые видны в микроскоп (часто под термином "хромосомы" понимают хроматин), хроматин содержит до 10% РНК. Две третьих оставшейся массы хроматина - белки, одна третья - ДНК. Половина всех белков хроматина - гистоны, для которых характерно высокое содержание лизина и (или) аргинина; это придает им щелочной характер и способность взаимодействовать с кислотными группами ДНК. Рибосомы - субклеточные частицы (см. рис. Стр.117) с молекулярной массой 4,5млн. Да, и коэффициентом седиментации 80S. Они состоят из двух субъединиц большой (коэффициент седиментации 60) и малой (40S). Большая 6ОS-субъединица составлена из двух молекул РНК (23S и 5S) и примерно 34 молекул белка. Меньшая 40S-субъединица содержит одну молекулу РНК (18S), состоящую из 1700 нуклеотидов, и 21 молекулу белка. Рибосома в целом функционирует как устройство для синтеза белков. Общее, содержание ДНК и РНК в клетках зависит от их функционального состояния. Например, в сперматозоидах количество ДНК достигает 60% (в пересчете на сухую массу клеток), в большинстве клеток -1-10, а в мышцах - около 0,2%. Доказано, что количественное содержание ДНК в клетках одного и того же организма отличается удивительным постоянством, однако различается для клеток разных организмов. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в печени, поджелудочной железе, эмбрионах и других тканях, активно синтезирующих белок, составляет от 4 до 10. В тканях с умеренным синтезом белка это соотношение колеблется от 0,3 до 2,5. Особое место занимают вирусы. У них в качестве генетического материала может быть либо ДНК (ДНК - вирусы), либо РНК (РНК - вирусы). В клетках бактерий, не имеющих ядра (прокариоты), молекула ДНК (хромосома) находится в специальной зоне цитоплазмы - нуклеозиде. В клетках, имеющих ядро (эукариоты), ДНК распределена между ядром, где она входит в состав хромосом (хроматин) и ядрышка, внеядерными органоидами (митохондриями и хлоропластами). Примерно 1-3% ДНК клетки приходится на внеядерную ДНК, остальное сосредоточено в ядре. Значит, наследственные свойства характерны не только для ядра, но и для митохондрий и хлоропластов клеток. РНК в отличие от ДНК распределена по клетке более равномерно, это говорит о том, что функции РНК более многообразны. В клетках высших организмов около 11% всей РНК находится в ядре, около 15% - в митохондриях, 50% - в рибосомах, и 24% - в гиалоплазме. 4.2. Химический состав и строение нуклеиновых кислот Нуклеиновыми кислотами называются высокомолекулярные соединения (полинуклеотиды), состоящие из остатков мононуклеотидов. Мононуклеотиды (нуклеотиды) построены из трех компонентов: азотистого основания (пиримидинового или пуринового), пентозы (рибоза или дезоксирибоза) и одной или нескольких фосфатных групп. В составе нуклеотидов выделяют более простой структурный компонент - нуклеозиды, состоящие из азотистого основания и пентозы. Азотистые основания делят на главные и редкие (минорные). К главным пуриновым основаниям относятся аденин и гуанин; к пиримидиновым - цитозин, урацил и тимин. В ДНК входят аденин, гуанин, цитозин, и тимин, а в РНК - все те же основания, кроме тимина, который заменен урацилом. Главные азотистые основания имеют следующее строение: 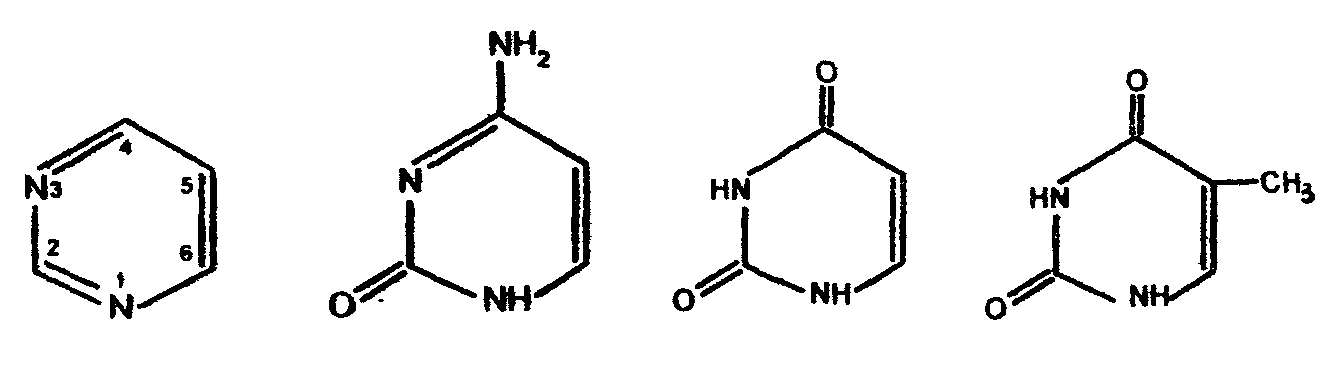
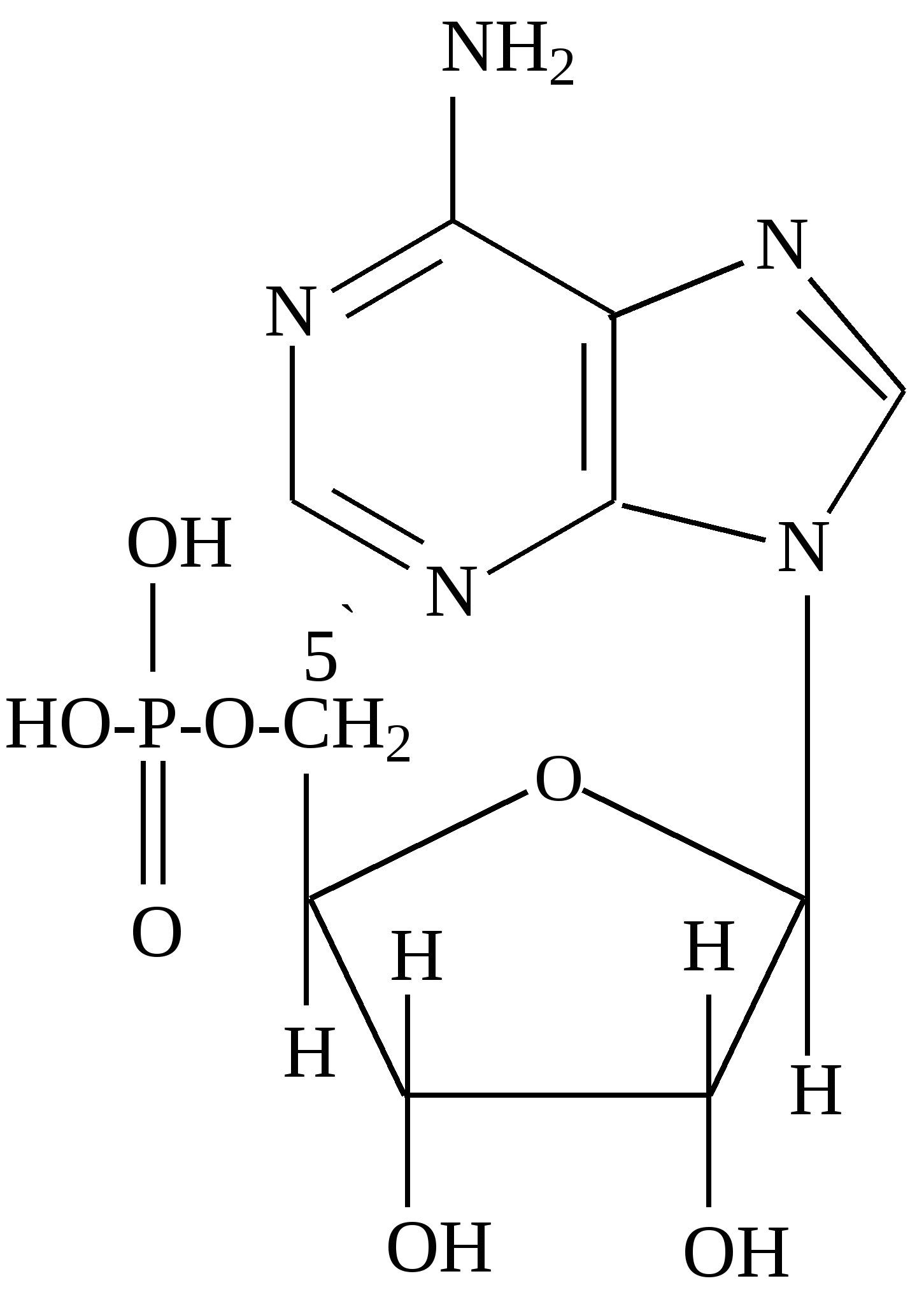
Минорные основания встречаются главным образом в тРНК. К ним относятся дополнительно метилированные пуриновые и пиримидиновые основания. В нуклеозидах атом углерода пентозы С-1´ связан с первым атомом азота (N-1) пиримидинового или с девятым атомом азота (N-9) пуринового основания. Пентоза в рибонуклеозидах - рибоэа, а в дезоксирибонуклеозидах - дезоксирибоза. Основные рибонуклеозиды - аденозин, гуанозин, цитидин и уридин. Основные дезоксирибонуклеозиды - дезоксиаденозин, дезоксигуаноэин, дезоксицитидин и дезокситимидин. Нуклеотиды представляют собой фосфорные эфиры нуклеозидов - нуклеозидфосфаты, образующиеся в результате этерификации гидроксильной группы при пятом атоме углерода пентозы (С - 5'). Удлинение фосфатного конца мононуклеотида за счет присоединения дополнительных фосфатов приводит к образованию нуклеозидполифосфатов. Ниже приведены примеры нуклеозидов, нуклеотидов и нуклеозидполифосфата:
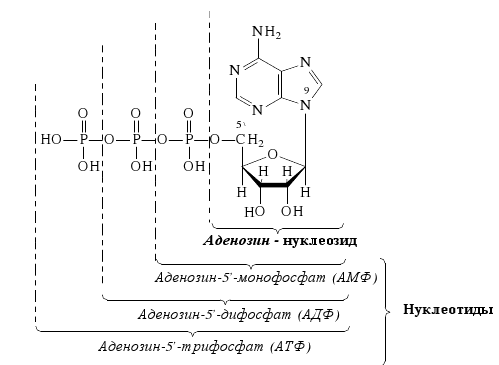 Чаще всего в клетках встречаются нуклезиддифосфаты и нуклеозидтрифосфаты. Мономерами РНК являются рибонуклеозидмонофосфаты, a мономерами ДНК – дезоксирибонуклеозидмонофосфаты. Ниже приводятся названия и сокращенные обозначения нуклеозидфосфатов:
Нуклеозидмонофосфаты кроме того называют следующим образом: АМФ-адениловая кислота, ГМФ-гуаниловая, ЦМФ-цитодиловая и УМФ-уридиловая кислота. В случае дезоксирибонуклеозид-монофосфатов добавляется приставка «дезокси», а вместо уридиловой кислоты в составе ДНК встречается дезокситимидиловая (дТМФ). Нуклеозидди- и трифосфаты содержат фосфодиэфирные высокоэнергетические связи, т.е. связи, при гидролизе которых освобождается значительное количество энергии (около 50 кДж/моль). Поэтому эти соединения участвуют в реакциях синтеза биологических веществ как источник энергии. Особенно уникальную роль в превращениях энергии в организме выполняет аденозинтрифосфат (АТФ). 4.3. Уровни структурной организации нуклеиновых кислот Нуклеиновые кислоты имеют первичную, вторичную и третичную структуру. Первичная структура нуклеиновых кислот Первичной структурой нуклеиновых кислот называют порядок чередования нуклеотидов (нуклеозидионофосфатов) в полинуклеотидной цепи. Мономеры в молекулах нуклеиновых кислот соединены сложноэфирной связью, образованной 5´- гидроксильной группой фосфатного остатка одного мононуклеотида и 3´ - гидроксильной группой пентозного остатка другого мононуклеотида (3', 5' - фосфодиэфирная связь). Ниже приведены фрагменты первичной структуры ДНК и РНК: 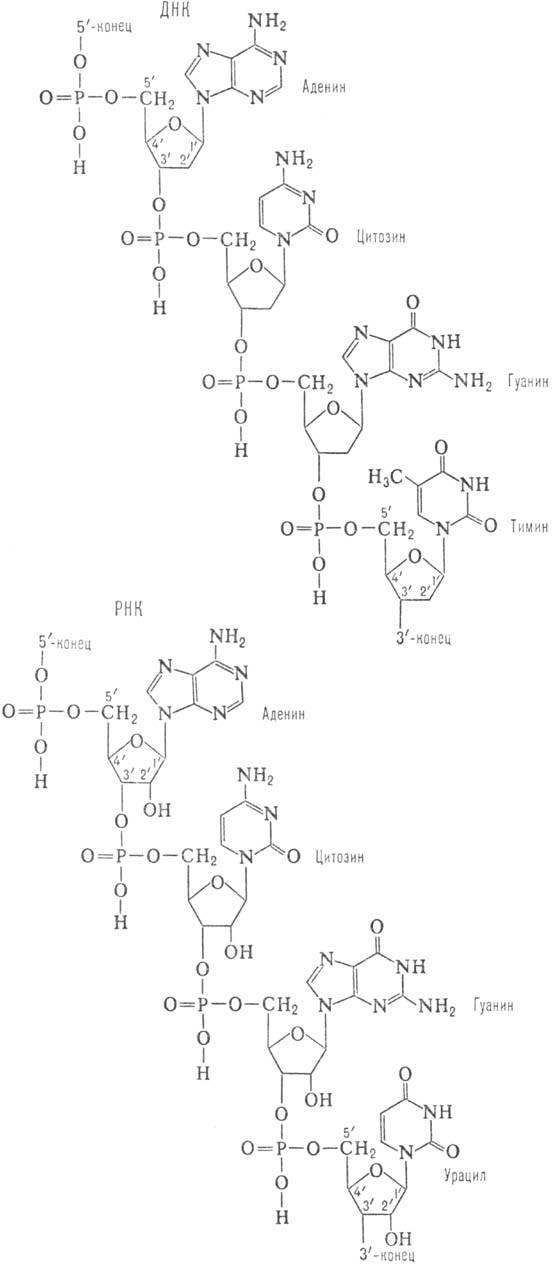 Линейные цепи ДНК и РНК, длина которых зависит от числа входящих в цепь нуклеотидов, имеют два конца: на одном имеется 5'- фосфатная ОН- группа (5'-конец), на другом - свободная З'-ОН- группа (3'-конец). Разные нуклеиновые кислоты отличаются друг от друга фактически чередованием азотистых оснований, так как пентозофосфатные части у всех мономеров одинаковы. Для краткого изображения первичной структуры нуклеиновых кислот пользуются однобуквенными символами азотистых оснований. Первичная структура РНК может быть представлена такой записью: (5´) А - У - А - А - Г - У - Ц - У-… (3-). Запись структуры ДНК отличается приставкой «д» (дезокси) и вместо символа «У» встречается символ «Т» : (5´) д (А - Т - А - Ц - Г - Т - А -...) (3'). Принимая средний молекулярный вес нуклеотидного остатка равным 330, легко подсчитать коэффициенты поликонденсации мономеров в молекулах нуклеиновых кислот. Учитывая, что молекулярная масса ДНК больше, чем РНК, для первых коэффициент поликонденсации всегда выше, чем для вторых. Генетический «текст» ДНК составлен с помощью кодовых «слов» - триплетов нуклеотидов (кодогенов). Участки ДНК, содержащие информацию о первичной структуре всех типов РНК, называют структурными генами. Порядок чередования нуклеотидов в РНК тот же, что и в копируемом участке ДНК, с той лишь разницей, что РНК состоит из рибонуклеотидов и вместо тимина в РНК - урацил. Первичная структура мРНК (набор кодонов) скопирована с участка ДНК, содержащего информацию о первичной структуре белковой молекулы (набор кодогенов). Первичная структура остальных типов РНК (тРНК, рРНК, редкие РНК) является окончательной копией генетической программы соответствующих структурных генов ДНК. В клетках высших организмов имеется 3 вида рРНК, различающихся по молекулярной массе (см.табл. 5), несколько десятков видов тРНК и огромное количество видов мРНК - не меньше, чем число разных белков в организме. Первичная структура нуклеиновых кислот определяет высшие уровни их организации - вторичную и третичную структуру. Вторичная структура ДНК Изучая состав ДНК, Чаргафф в 1949 г. установил важные закономерности, касающиеся отдельных оснований ДНК. Они помогли раскрыть вторичную структуру ДНК. Эти закономерности называют правилами Чаргаффа: 1) сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклеотидов, т.е. А+ГУЦ+Т = 1; 2) содержание аденина равно содержанию тимина: А=Т; 3) содержание гуанина равно содержанию цитозина: Г=Ц; 4) количество аденина и цитозина равно количеству гуанина и тимина: А+Ц=Г+Т; 5) изменчивы только суммы А+Т и Г+Ц. Если А+Т>Г+Ц, то это AT - тип ДНК; если Г+Ц>А+Т, то это ГЦ-тип ДНК. Эти правила говорят о том, что при построении ДНК должно соблюдаться довольно строгое соответствие не пуриновых и пиримидиновых оснований вообще, а конкретно тимина с аденином и цитозина с гуанином. На основании правил Чаргаффа и рентгеноструктурных исследований Дж. Уотсон и Ф. Крик ( Великобритания ) предложили модель строения ДНК (1953), основные черты которой сформулированы ниже. 1. Молекула ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно и на всем протяжении связанных друг с другом водородными связями. 2. Водородные связи между цепями образуются за счет специфического комплементарного взаимодействия аденинового остатка одной цепи с тиминовым остатком другой цепи (пара А - Т) и гуанинового остатка одной цепи с цитозиновым остатком другой цепи (пара Г - Ц): Основания А и Т, Ц и Г комплементарны друг другу в том смысле, что между ними легче возникают водородные связи, чем при других сочетаниях (например, А и Г, Ц и А и т.д.). 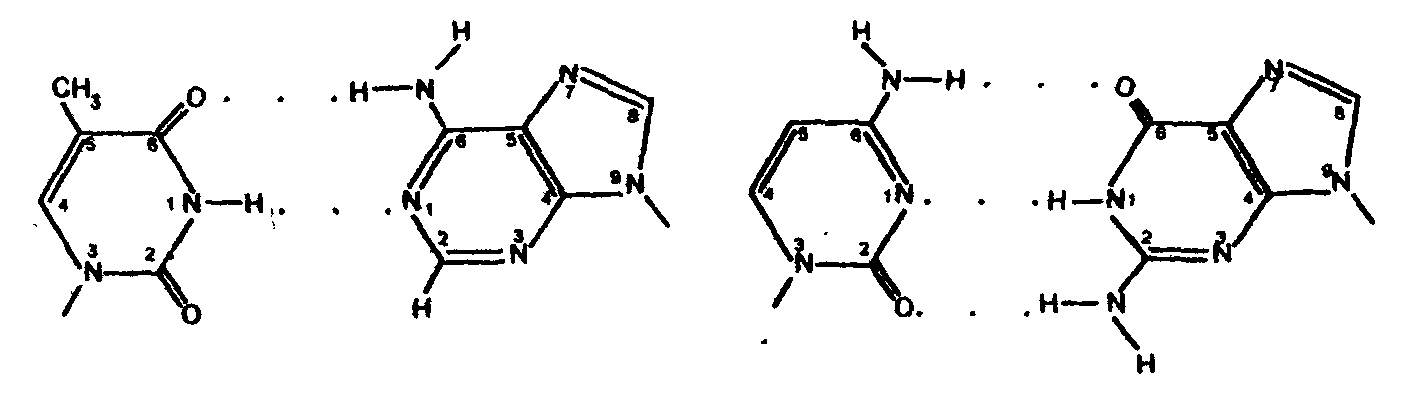
3. Первичная структура одной цепи молекулы ДНК комплементарна первичной структуре другой цепи. Это можно показать следующей схемой: 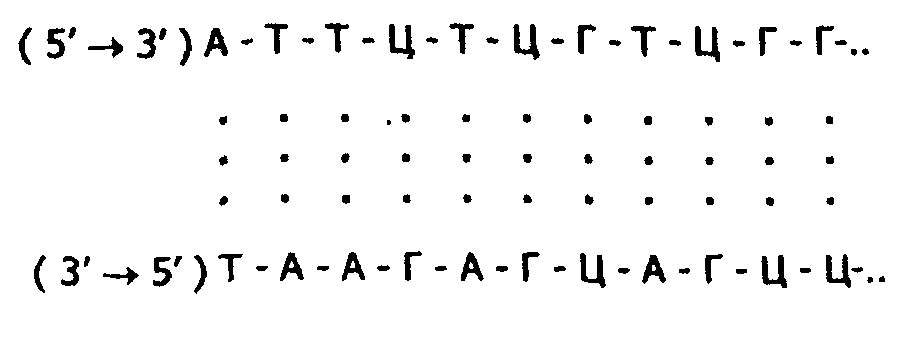 Таким образом, если известна первичная структура одной цепи, легко может быть написана, исходя из правил комплементарности, первичная структура другой цепи. 4. Обе цепи закручены в спираль, имеющую общую ось; цепи могут быть разъединены только путем раскручивания. Пуриновые и пиримидиновые основания обращены внутрь спирали; их плоскости перпендикулярны оси спирали и параллельны друг другу, образуя таким образом стопку оснований. Между основаниями в этой стопке возникают гидрофобные взаимодействия, которые стабилизируют двойную спираль даже больше, чем водородные связи между цепями. Пентозофосфатные части располагаются по периферии, образуя ковалентный остов спирали. По Уотсону и Крику: расстояние между витками (шаг спирали) равно 3,4 нм. На этом участке укладывается 10 нуклеотидных остатков, размер одного нуклеотида 0,34 нм, диаметр биспиральной молекулы равен 1,8 нм. 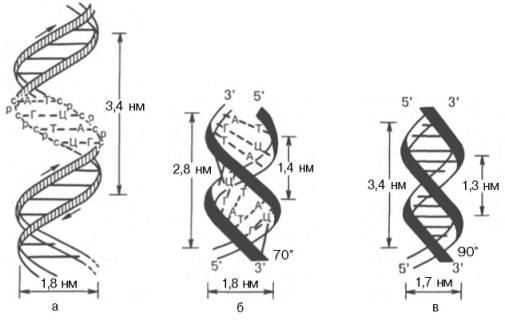 Рис. Схематическое изображение двойной спирали ДНК: а - по Уотсону и Крику; б - А-форма ДНК; в - В-форма ДНК (С - остаток дезоксирибозы; Р - остаток фосфорной кислоты) В настоящее время методами рентгеноструктурного анализа установлено существование четырех форм ДНК: А, В, С, Т. Простейшие из них А и В. У формы А наблюдается смещение пар оснований от оси молекулы к периферии, поэтому уменьшается шаг спирали до 2,8 нм, и в одном витке 11 мононуклеотидов вместо 10. В-форма приближается к модели Уотсона и Крика: шаг спирали 3,4 нм, диаметр биспирали - 1,7 нм . Между А- и В- формами ДНК осуществляются взаимопереходы, которые могут быть вызваны растворителями или белками. В этих переходах, очевидно, заключен определенный биологический смысл. Предполагается, что в А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК на матрице ДНК), а в В-форме - роль матрицы в процессе репликации (синтез ДНК на матрице родительской ДНК). Установлено, что в клетках ряда бактерий и вирусов ДНК существует в виде одноцепочечной молекулы, вторичная структура которой представляет собой сочетание неспирализованных участков с участками спирали, образованной за счет взаимодействия комплементарных друг другу оснований. Описанная структура ДНК позволяет объяснить молекулярный механизм функциональных биологических явлений, таких как самовоспроизведение организмов, наследственность, изменчивость. Поэтому. 1953 г., когда Ф. Крик и Дж. Уотсон разработали модель строения ДНК, принято считать годом рождения молекулярной биологии. Вторичная структура РНК Молекулы РНК в отличие от ДНК построены из одной полинуклеотидной цепи. Однако в этой цепи (для рРНК и мРНК) имеются комплементарные друг другу участки, которые могут взаимодействовать, образуя двойные спирали. При этом соединяются водородными связями нуклеотидные пары А-У и Г-Ц. Такие спирализованные участки (их называют шпильками) обычно содержат небольшое количество нуклеотидных пар (до 20-30) и чередуются с неспирализованными участками. Характерную вторичную структуру имеют тРНК. Они содержат четыре спирализованных участка и три (четыре) одноцепочные петли. При изображении такой структуры на плоскости получается фигура, называемая «клеверным листом» (рис. справа). 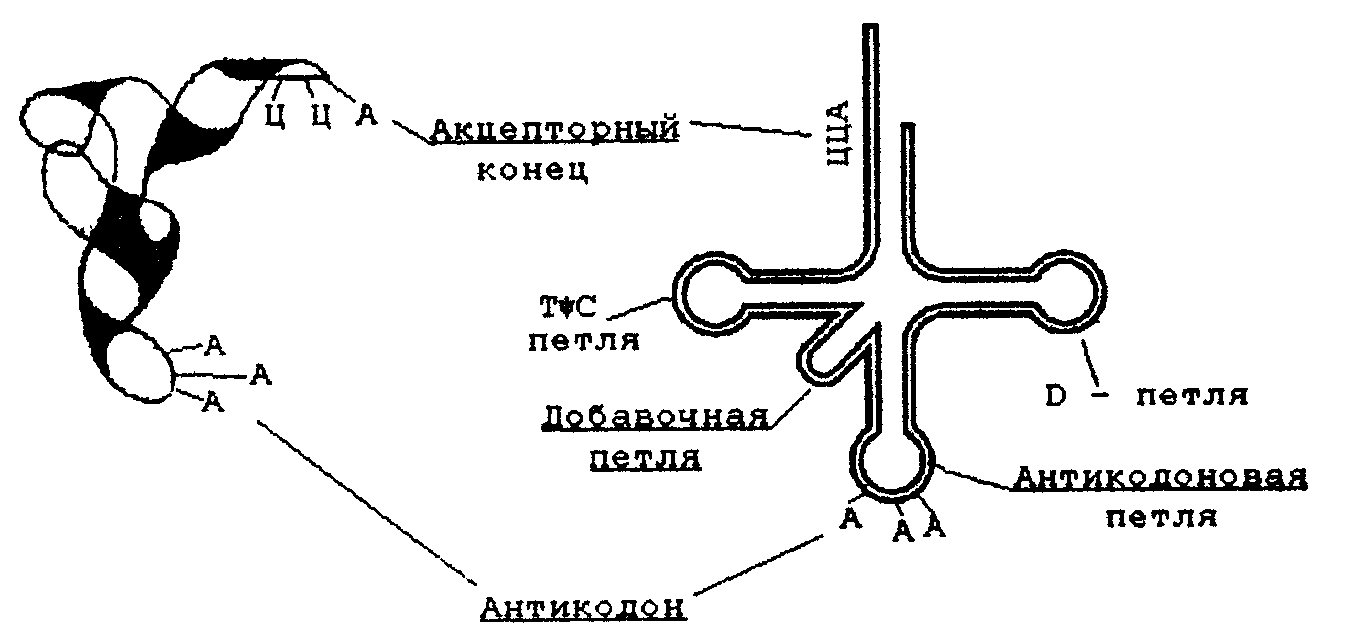 Рис.. Вторичная (справа) и третичная (слева) структура тРНК Все несколько десятков разных тРНК клетки имеют общий план пространственной структуры, но различаются в деталях. В тРНК выделяют следующие структурные участки. 1. Акцепторный конец - во всех типах тРНК имеет состав ЦЦА. К гидроксилу З'-ОН аденозина карбоксильной группой присоединяется аминокислота, которую данная тРНК доставляет к рибосомам, где происходит синтез белка. 2. Антикодоновая петля - содержит специфический для каждой тРНК триплет нуклеотидов (антикодоны). Антикодон комплементарен кодону мРНК. Кодон-антикодоновое взаимодействие определяет порядок чередования аминокислот в белковой молекуле при синтезе ее на рибосомах. 3. Псевдоуридиловая петля (Г,С) - участвует в связывании тРНК с рибосомой. 4. Дигидроуридиловая (D) петля необходима для связывания с ферментом аминоацил-тРНК-синтетзой, которая участвует в узнавании аминокислотой своей тРНК. 5. Добавочная петля - разная у разных тРНК. Третичная структура РНК и ДНК Пространственная конфигурация спирализованной полинуклеотидной цепи (третичная структура) достаточно полно выяснена для молекул РНК. Установлено, что нативные молекулы тРНК имеют примерно одинаковую третичную структуру, которая отличается от плоской структуры «клеверного листа» (вторичная структура) большей компактностью за счет складывания различных частей молекулы (см. рис выше). Для рРНК и мРНК возможно существование, а зависимости от концентрации солей и температуры, трех видов третичной структуры (рис. ниже). Первый - рыхлый беспорядочный клубок или распрямленная цепь (при повышении температуры и отсутствии солей). Второй вариант - компактный клубок с двуспиральными участками (высокая ионная сила, комнатная температура). Третий вид - компактная палочка с упорядочение ориентированными двуспиральными участками (низкая ионная сила, комнатная температура). Все три типа третичной структуры РНК связаны взаимными переходами. Третичная структура ДНК зависит от того, сколько цепочек полинуклеотидов (одна или две) в ДНК. В ряде вирусов обнаружены одноцепочечные ДНК линейной и кольцевой формы. Двуцепочечные спиралевидные молекулы ДНК также могут существовать в линейной и кольцевой форме; образование последней вызвано ковалентным соединением их открытых концов. 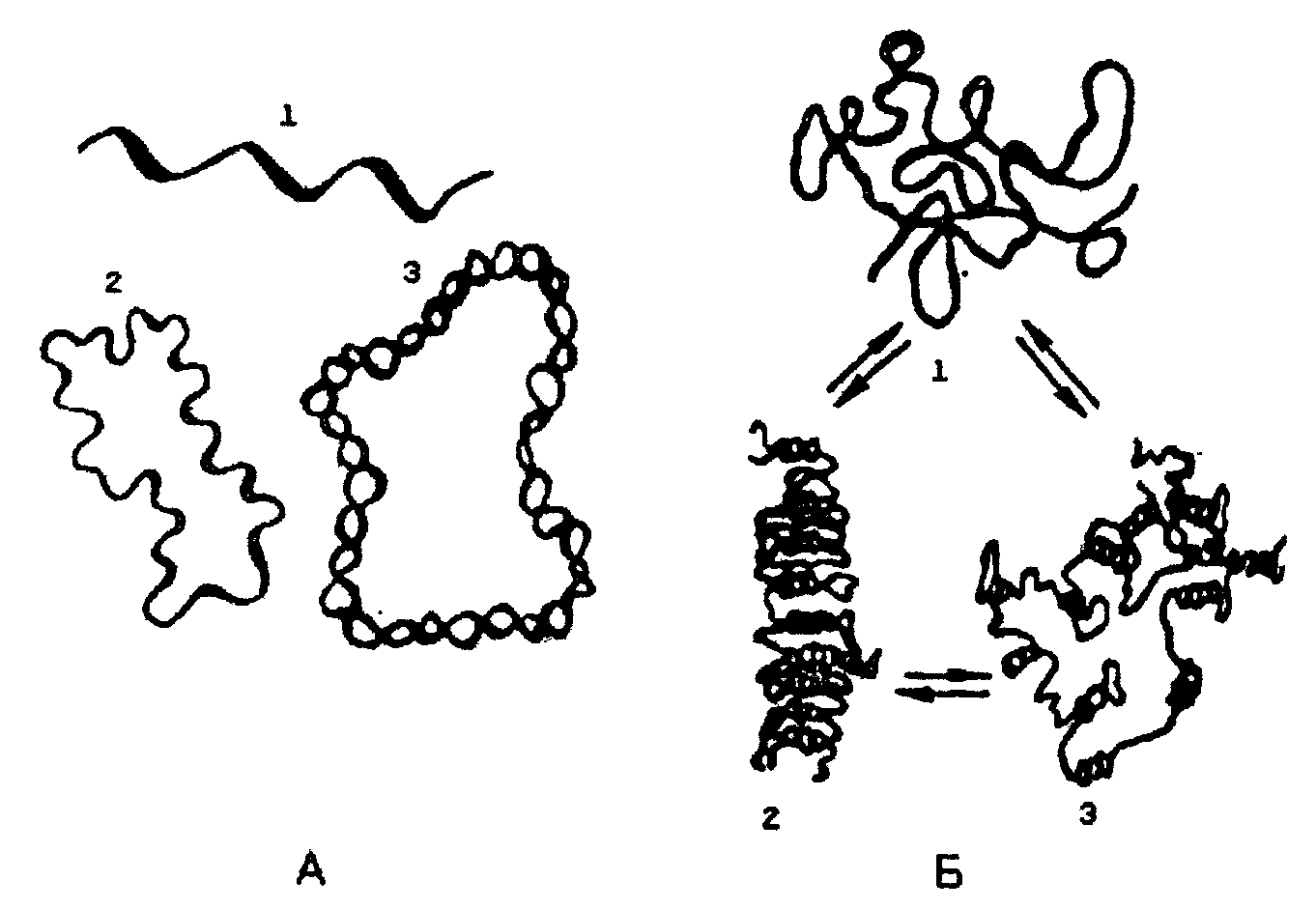 Рис. Третичная структура: А - ДНК: 1 - линейная одноцепочечная бактериофаг ФХ174 (и др. вирусов); 2 - кольцевая одноцепочечная ДНК вирусов и митохондрий; 3 - кольцевая двойная спираль ДНК; Б - РНК: 1 - рыхлый клубок или распрямленная цепь; 2 - компактная палочка; 3 - компактный клубок Кроме того, полагают, что биспиральные молекулы ДНК существуют в хромосомах в виде вторично спирализованных фрагментов, связанных друг с другом (суперспираль). Поэтому молекулярный вес нативной ДНК достигает нескольких сотен миллионов. Следовательно, молекулы с молекулярной массой 10.000.000 являются субъединицами более крупных молекулярных образований (третичная структура). Именно суперспирализация обеспечивает экономную упаковку огромной молекулы ДНК в хромосоме: вместо 8 см длины, которую она могла бы иметь в вытянутой форме, она занимает всего 5 нм. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||