Ответы к экзамену По биохимии 1 структура и функции белков 3 ферменты 10 нуклеиновые кислоты и нуклеотиды 21
 Скачать 29.52 Mb. Скачать 29.52 Mb.
|

ОБМЕН УГЛЕВОДОВВопрос №48 Общая характеристика, функции и классификация углеводов. Суточная потребность, расщепление до моносахаров в ЖКТ. Примеры нарушения переваривания. Углеводы – производные многоатомных спиртов, альдегидов или кетонов. Г 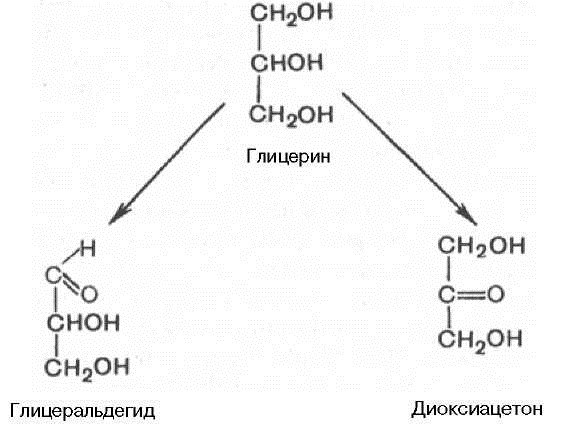 лицерол – спирт с наименьшим количеством атомов, из которых можно образовать углевод. лицерол – спирт с наименьшим количеством атомов, из которых можно образовать углевод.Если есть альдегидная группа – глицероальдегид (ГАД) Если есть кетогруппа – диоксиацетон (ДАЦ) Основная функция углеводов: Углеводы – основной источник энергии организма, т.к. до 80% энергозатрат обеспечивается углеводами. Мозг использует только углеводы (глюкозу). Суточная потребность углеводов – 400-500 г (содержится в картофеле, крупах, конфетах, фруктах). Углеводы:
Если остатки одного моносахарида – гомополисахарид (функция – запасающая) Если остатки двух и более – гетерополисахарид (функция – строительная) Гомополисахариды:
Углеводы распадаются в 3 стадии с целью увеличения энергии 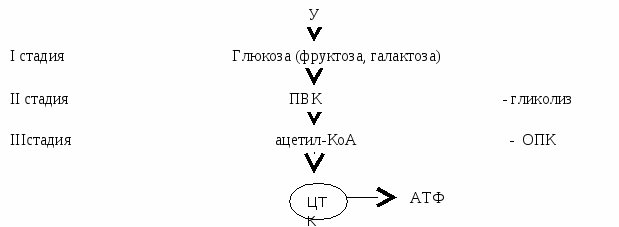 I стадия – переваривание (в ЖКТ) В 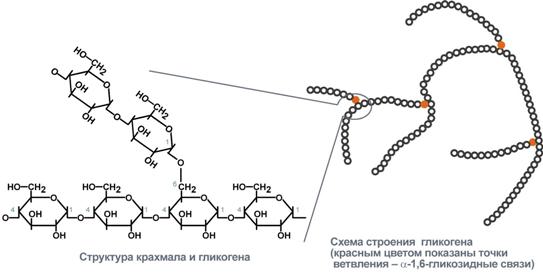 крахмале глюкозные остатки в линейных участках соединены 1→4-гликозидными связями, а точках ветвления – 1→6. крахмале глюкозные остатки в линейных участках соединены 1→4-гликозидными связями, а точках ветвления – 1→6. Особенности:
А) полостное пищеварение – под действием ферментов поджелудочной железы Б) пристеночное – под действием ферментов клеток кишечника 1 – пищеварение в ротовой полости Под действием фермента околоушной слюной железы – α-амилаза – в углеводах расщепляются линейные связи (1→4-гликозидные). Т.к. пища находится в ротовой полости недолго, то полисахариды (крахмал) расщепляются до фрагментов – декстринов. 2 – углеводы в желудке не перевариваются 3А (полостное пищеварение):– в кишечнике на декстрины действуют 3 фермента поджелудочной железы
В результате полостного пищеварения образуется мальтоза (глюкоза-1→4-глюкоза) и изомальтоза (глюкоза-1→6-глюкоза) 3Б – Пристеночное пищеварение: Дисахариды расщепляются ферментами клеток кишечника. Здесь расщепляются не только мальтоза и изомальтоза, но и сахароза и лактоза, которые расщепляются только в пристеночном пищеварении. Клетки кишечника вырабатывают ферменты, расщепляющие дисахариды на составные компоненты (ферменты мальтаза, изомальтаза, лактаза, сахараза). Около 80% фруктозы в кишечнике расщепляется до глюкозы. Т.е. глюкоза – основной моносахарид, образующийся в кишечнике при переваривании. Возможные нарушения переваривания, связанные с врожденной (жители Азии) или временной (дети) недостаточностью лактазы. II стадия – всасывание: Всасывание глюкозы может идти 2 путями (в зависимости от состояния – голодный или сытый). О состоянии будем судить по концентрации глюкозы в крови:
Таким образом, концентрация глюкозы в крови 3,5-5,5 Мм/лявляется оптимальной для работы мозга, т.к. мозг использует столько глюкозы, сколько ее в крови. Голодный: После приема пищи концентрация глюкозы в кишечнике выше, чем в крови. Следовательно, она всасывается по градиенту облегченной диффузией с белком-переносчиком. Белки-перенсочики = ГЛЮТ (в зависимости от ткани): ГЛЮТ-1 – мозг, плацента ГЛЮТ-2 – поставщик глюкозы в кровь (кишечник, печень) ГЛЮТ-3 – остальные ткани ГЛЮТ-4 – единственный переносчик в инсулин-зависимые ткани (мышцы, жировая ткань) ГЛЮТ-5 – неинсулин-зависимые ткани Инсулин-зависимые ткани – ткани, в которые глюкоза поступает только в присутствии инсулина. С 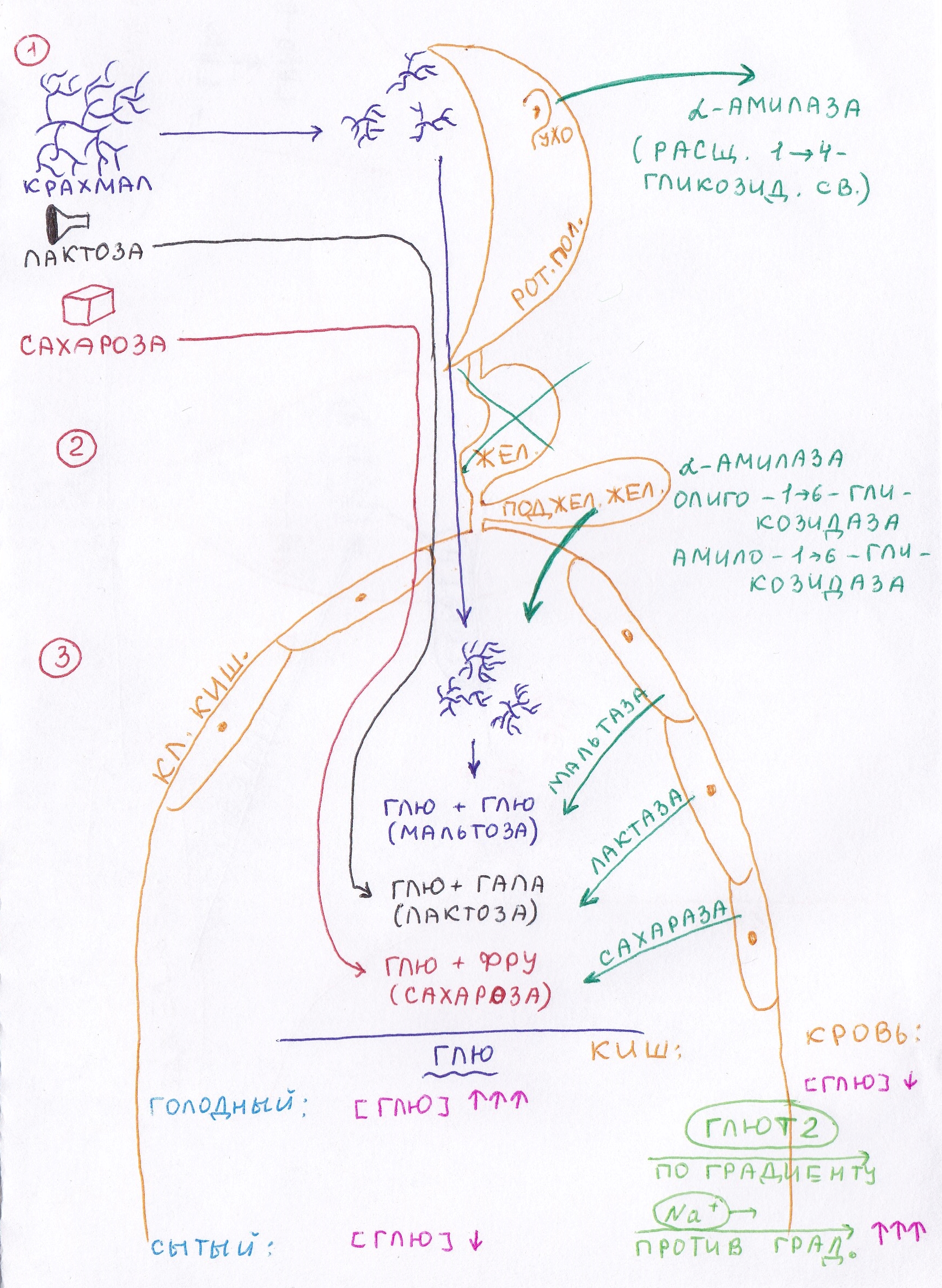 ытый: ытый:Концентрация глюкозы в крови выше, чем в кишечнике. Следовательно, глюкоза всасывается против градиента активным транспортом (симпорт с Na+). Абсорбтивный период – после принятия пищи. Постабсорбтивный период – между приемами пищи. Переваривание углеводов занимает 30 мин – 1,5 часа. Основная реакция, которую активирует инсулин – фосфорилирование глюкозы. Глюкоза → Глюкоза-6Ф Глюкоза-6Ф не может выйти из клеток в кровь и используется ими. Следовательно, основное вещество, используемое клетками – не глюкоза, а глюкоза-6Ф. 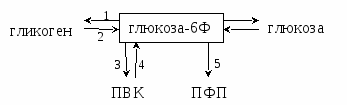 Вопрос №51 Гликогенная функция печени, биосинтез и мобилизация гликогена, зависимость от ритма питания. Гормональная регуляция.
Гликоген запасается в печени (6% от массы печени) -120 г, и мышцах (1% от массы мышц). В печени распад и синтез гликогена направлены на поддержание нормальной концентрации глюкозы в крови. В мышцах – на обеспечение энергией только мышц. Синтез гликогена активируется инсулином. Распад синтезируется в печени – глюкагоном и адреналином, в мышцах – только адреналином. Гликоген запасается в виде гранул (чтобы не повышалось осмотическое давление в клетках). На синтез гликогена требуется 2 АТФ. Гексокина - в мышцах, глюкокиназа - в печени поджелудочной. 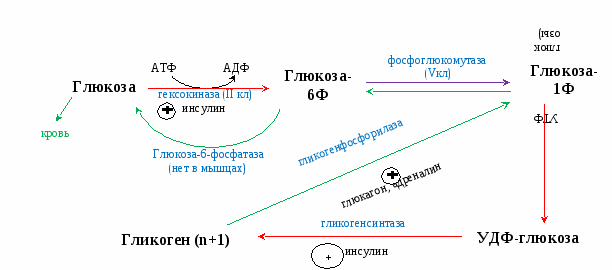  Распад гликогена активируется между приемами пищи, при длительной физической нагрузке или длительном голодании, когда концентрация глюкозы в крови понижена. Выделяется глюкагон и активирует гликогенфосфорилазу. В мышцах синтез гликогена идет точно так же, а распад идет только до глюкозы-6Ф (нет фермента, дефосфорилирующего глюкозу – глюкоза-6-фосфатаза). Глюкоза-6Ф остается в мышцах, используется как источник энергии. Активируется процесс только адреналином. Вопрос №52 Гормональная регуляция обмена гликогена в печени и мышцах. Наследственные нарушения обмена гликогена. Гликогеновые болезни —наследственных нарушений, снижение или отсутствие активности ферментов, катализирующих реакции синтеза или распада гликогена, либо нарушение регуляции. Гликогенозы — заболевания, обусловленные дефектом ферментов, участвующих в распаде гликогена. Они проявляются необычной структурой гликогена или избыточным накоплением в печени, сердечной или скелетных мышцах, почках, лёгких и других органах. Гликогенозы: печёночные и мышечные. Печёночные формы гликогенозов ведут к нарушению использования гликогена для поддержания уровня глюкозы в крови. Поэтому общий симптом — гипогликемии в постабсорбтивный период. Болезнь Гирке — наследственный дефект глюкозо-6-фосфатазы. Клетки печени и извитых канальцев почек заполнены гликогеном, печень и селезенка увеличены, у больных опухлое лицо - «лицо китайской куклы». Болезнь проявляется гипогликемией, гиперурикемией, ацидоз. 1). В гепатоцитах: ↑ глюкозо-6-ф → ↑ ПВК, ↑ лактат (ацидоз), ↑ рибозо-5-ф → ↑ пуринов→ ↑ мочевая кислота, 2). В крови: ↓ глюкоза → ↓ инсулин/глюкагон→: а) ↑ липолиз жировой ткани → ↑ ЖК в крови, б) ↓ ЛПЛ жировой ткани → ↑ ТАГ. Болезнь Кори - накапливается разветвленный гликоген, дефектен деветвящий фермент. Гликогенолиз в незначительном объёме. Лактоацидоз и гиперурикемия не отмечаются. Мышечные формы - нарушение в энергоснабжении скелетных мышц (проявляются при физических нагрузках и сопровождаются болями и судорогами в мышцах, слабостью и быстрой утомляемостью). Болезнь МакАрдла —отсутствует активность гликогенфосфорилазы. Накопление в мышцах гликогена аномальной структуры. Агликогеноз— дефект гликогенсинтазы. В печени и других тканях очень низкое содержание гликогена. Гипогликемией в постабсорбтивном периоде. Судороги, особенно по утрам. Больные дети в частом кормлении. Вопрос №54 Гликолиз. Аэробный путь расщепления углеводов. Энергетика процесса. Челночные механизмы транспорта водорода из цитоплазмы в митохондрию. Гликолиз Гликолиз – специфический путь катаболизма глюкозы до ПВК. И  меет энергетическую значимость (обеспечивает 70% потребления энергии). меет энергетическую значимость (обеспечивает 70% потребления энергии).Локализация – в цитоплазме клеток всех органов. Сущность: Линейный дихотомический процесс Все промежуточные продукты гликолиза – в фосфорилированной форме. Донор фосфора - АТФ. В процессе – 10 реакций и 2 этапа: 1 – до образования двух триоз (ДАФ, ГАФ), затрачивается 2 АТФ на фосфорилирование в 1 и 3 реакциях. 2 – период ОВ-реакций, т.к. есть одна реакция окисления, за счет которой будет синтезироваться АТФ путем окислительного фосфорилирования (реакция №6). В 7 и 10 реакциях синтезируется 2 АТФ путем субстратного фосфорилирования. 1, 3, 10 реакции – необратимые, и являются регуляторными. Регуляторный фермент – фермент 3 реакции – фосфофруктокиназа (ФФК) – аллостерический, определяет скорость гликолиза. Энергетический выход гликолиза – 6 (8) АТФ. Образованное ПВК поступает в ОПК (ОДПВК и ЦТК). Т.к. из глюкозы получается 2 триозы (диоксиацетонфосфат, глицероальдегидфосфат), то ДАФ → ГАФ и второй этап проходит еще раз. Т.е. при расщеплении 1 глюкозы первая стадия проходит один раз, а вторая – два. С 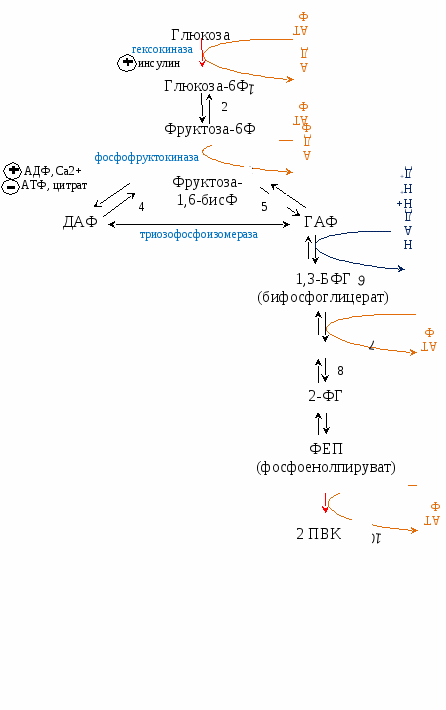 ледовательно, гликолиз – первый этап полного окисления глюкозы до СО2 и Н2О. ледовательно, гликолиз – первый этап полного окисления глюкозы до СО2 и Н2О.  3-ФГ 9 Подсчет энергии (суммарный выход гликолиза): ∑Е=ЕSф + EОф = 4 + 4 (6) = 8 (10) – 2 (на первом этапе) = 6 (8) АТФ ЕSф – субстратное фосфорилирование (7 и 10 реакции) – 2 АТФ. Т.к. 2 этап проходит дважды, то 2х2=4 АТФ. EОф – окислительное фосфорилирование –за счет 6 реакции. Если бы она протекала в митохондрии, то 2Н поступали бы в ЦПЭ на первом этапе (НАДН-ДГ). Но она протекает в цитоплазме, и поэтому нужно перенести 2Н из цитоплазмы в митохондрии. Для этого существуют челночные механизмы переноса водорода из цитоплазмы в митохондрии:
Глицеротрифосфатный челночный механизм: 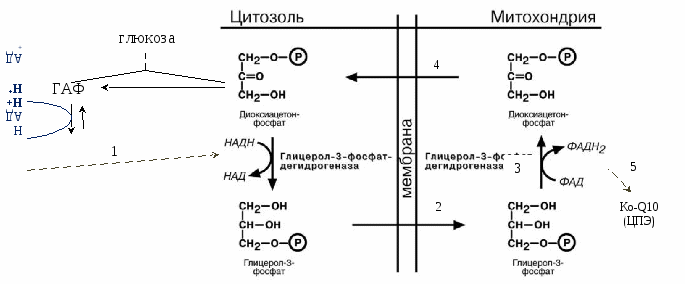 3
Следовательно, за 6 реакцию при помощи этого механизма синтезируется 2 АТФ. Т.к. 2 этап проходит дважды, то за 6 реакцию выделяется 2х2 АТФ (или 2х3 АТФ при использовании второго механизма). Подсчет энергии полного окисления глюкозы:  ВСЕГО: 6 (8) (гликолиз) + 2х3 (ОДПВК) + 2х12 (ЦТК) = 36 (38) АТФ Вопрос №55 Анаэробный гликолиз. Примеры, энергетика процесса. В  некоторых клетках (эритроциты, опухолевые клетки, кардиомиоциты при гипоксии) и органах (мышцы при интенсивной кратковременной работе) ПВК превращается не в ацетил-КоА, а восстанавливается до лактата (неправильно – анаэробный гликолиз). некоторых клетках (эритроциты, опухолевые клетки, кардиомиоциты при гипоксии) и органах (мышцы при интенсивной кратковременной работе) ПВК превращается не в ацетил-КоА, а восстанавливается до лактата (неправильно – анаэробный гликолиз). При этом энергетический выход составляет 2 АТФ (только субстратное фосфорилирование). 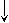 ∑Е = 2х2 = 4 – 2 (на первом этапе) = 2 АТФ За счет 6 реакции не считается, т.к. кофермент используется в 11 реакции. Вопрос №56 Пути обмена лактата в мышцах и печени. Вопрос №61 Глюконеогенез из молочной кислоты. Глюкозо-лактатный цикл. 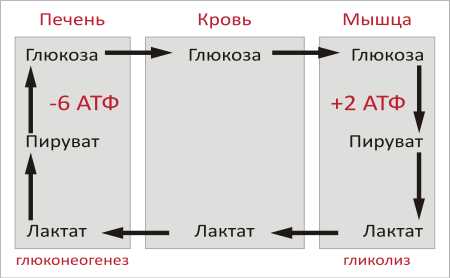 цикл Корипоказывает взаимосвязь между гликолизом в мышцах и ГНГ в печени. цикл Корипоказывает взаимосвязь между гликолизом в мышцах и ГНГ в печени.1 - обеспечивает утилизацию лактата; 2 - предотвращает накопление лактата и, как следствие этого, опасное снижение рН (лактоацидоз). Часть пирувата, образованного из лактата, окисляется печенью до СО2 и Н2О. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза. Цикл Кори:
Вопрос №57 Изменения объема глюкозы в печени (синтез и распад гликогена, гликолиз) в зависимости от ритма питания и мышечной активности. Роль гормонов в переключении процессов. 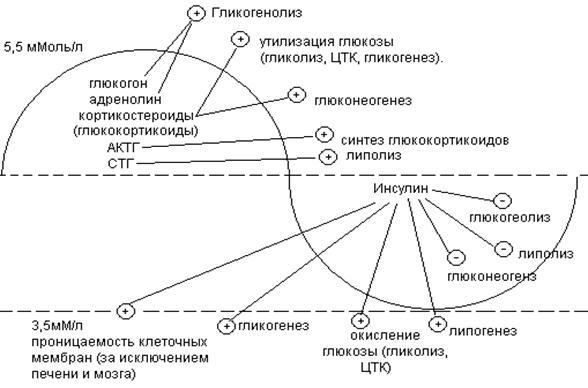 Вопрос №58 Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, жировая, печень. Э 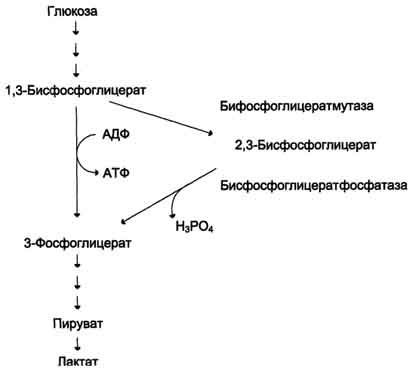 ритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути. ритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути. Лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа+, К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях. Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками - присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином. Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глутатиона. Мозг расходует свыше 20% всей энергии потребляемой организмом. Основная доля энергетической потребности мозга удовлетворяется посредством катаболизма глюкозы в ЦТК. Кетоновые тела могут использоваться как энергетический субстрат только при длительном голодании как дополнительный источник энергии. Повышенная утилизация кетоновых тел наблюдается у плода и в раннем детском возрасте, а у взрослых - при стрессе и при сахарном диабете, гипертиреозе, но покрывается не более 20% потребности. В качестве дополнительного источника энергии могут служить аминокислоты. Распад аминокислот осуществляется путем метаболизма, которую называют шунтом гамма-аминомаслянной кислоты (ГАМК). В случае увеличения расхода энергии и дефицита глюкозы увеличивается содержание ГАМК, которая является тормозным медиатором для нейронов и снижает функциональную активность в определенных структурах мозга. Этот процесс может рассматривается как регуляция по механизму отрицательной обратной связи: высокая церебральная активность, вызывает дефицит глюкозы, который запускает шунт ГАМК, в результате чего накапливается ГАМК, тормозящая активность нейронов. В норме использование шунта ограничено. Особенность энергообмена головного мозга заключается в том, что он практически не содержит запасов веществ используемых в качестве энергетического субстрата и постоянно нуждается в их поступлении через мозговой кровоток. 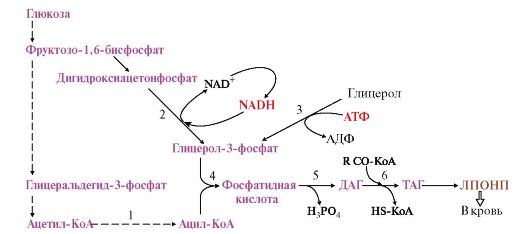 Синтез жиров в печени происходит в основном из продуктов катаболизма глюкозы. После еды образующийся в результате гликолиза избыток ацетил-КоА активно используется для синтеза жирных кислот (1). Глицерол-3-фосфат образуется двумя путями (2, 3). Синтез ТАГ происходит через образование фосфатидной кислоты (4), при дефосфорилировании которой образуется ДАГ (5). Следующая реакция ацилирования превращает его в ТАГ (6). ТАГ в составе ЛПОНП секретируются в кровь Синтез жиров в печени происходит в основном из продуктов катаболизма глюкозы. После еды образующийся в результате гликолиза избыток ацетил-КоА активно используется для синтеза жирных кислот (1). Глицерол-3-фосфат образуется двумя путями (2, 3). Синтез ТАГ происходит через образование фосфатидной кислоты (4), при дефосфорилировании которой образуется ДАГ (5). Следующая реакция ацилирования превращает его в ТАГ (6). ТАГ в составе ЛПОНП секретируются в кровьВопрос №63 Обмен пировиноградной кислоты в тканях. Он является конечным продуктом метаболизма глюкозы в процессе гликолиза. В условиях достаточного поступления кислорода превращается в ацетил-кофермент А. Пируват может быть превращён в анаплеротической реакции в оксалоацетат, который окисляется до углекислого газа и воды. При анаэробном дыхании в клетках пируват, полученный при гликолизе, преобразуется в лактат при помощи фермента ЛДГ и NADP. ПВК является «точкой пересечения» многих метаболических путей. Пируват может быть превращён обратно в глюкозу, в жирные кислоты или энергию через ацетил-КоА, в аланин. Пировиноградная кислота содержится во всех тканях и органах и, являясь связующим звеном обмена углеводов, жиров и белков, играет важную роль в обмене веществ. Концентрация пировиноградной кислоты в тканях изменяется при болезнях печени, некоторых формах нефрита, раке, авитаминозах, особенно при недостатке витамина В1. Нарушение обмена пировиноградной кислоты приводит к ацетонурии. Вопрос №59 Пентозофосфатный путь превращения глюкозы. Пентозофосфатный путь (ПФП) ПФП – альтернативный путь окисления глюкозы, но энергетического значения не имеет. Значение – поставляет важные компоненты:
Локализация - в цитоплазме клеток печени, эритроцитов, жировой ткани, молочной железы в период лактации Регуляторный фермент - глюкозо-6Ф-ДГ (при его дефиците развивается гемолитическая анемия). В ПФП различаю 2 пути:
Образуется НАДФН+Н+ (восстановленный) и рибулоза-5Ф (из нее образуется рибоза-5Ф).
Участвуют трансальдолазы и транскетолазы (кофермент ТДФ). Связан с гликолизом через ГАФ и фруктоза-6Ф. Р 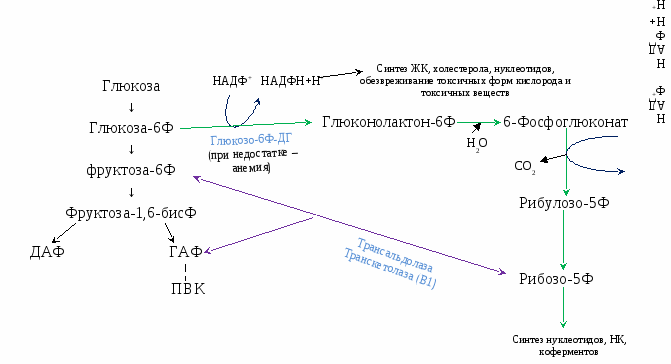 егуляторный фермент находится на 1 стадии. егуляторный фермент находится на 1 стадии.В зависимости от потребностей организма различают:
ПФП – Если организму требуется и НАДФН+Н+ и рибозо-5Ф (в печени), то протекают только окислительные реакции. ПФЦ – Если требуется только НАДФН+Н+ и мало рибозо-5Ф, то будут протекать и окислительные, и неокислительные реакции, которые превращают рибозу-5Ф в промежуточные продукты гликолиза (жировая ткань). ПФШ – Если нужно много рибозо-5Ф и мало НАДФН+Н+, то окислительный этап не протекает, а рибозо-5Ф образуется из промежуточных продуктов гликолиза (фруктозо-6Ф). Вопрос №60 Обмен лактозы и галактозы 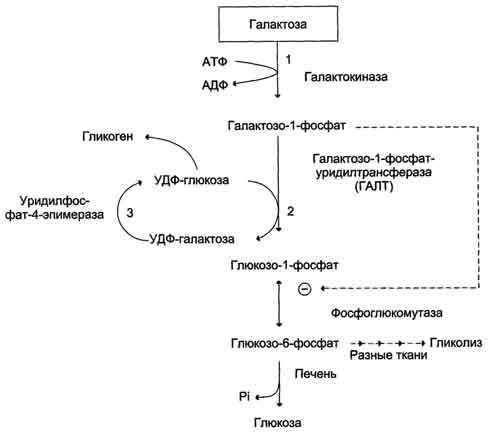 Вопрос №62 Глюконеогенез из аминоксилот и глицерола. Био значение. Глюкозо-аланиновый цикл. С  интез из аминокислот. В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма. Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты цитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных. Например, оксалоацетат, образующийся из Асп, является промежуточным продуктом как цитратногр цикла, так и глюконеогенеза. интез из аминокислот. В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма. Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты цитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных. Например, оксалоацетат, образующийся из Асп, является промежуточным продуктом как цитратногр цикла, так и глюконеогенеза.  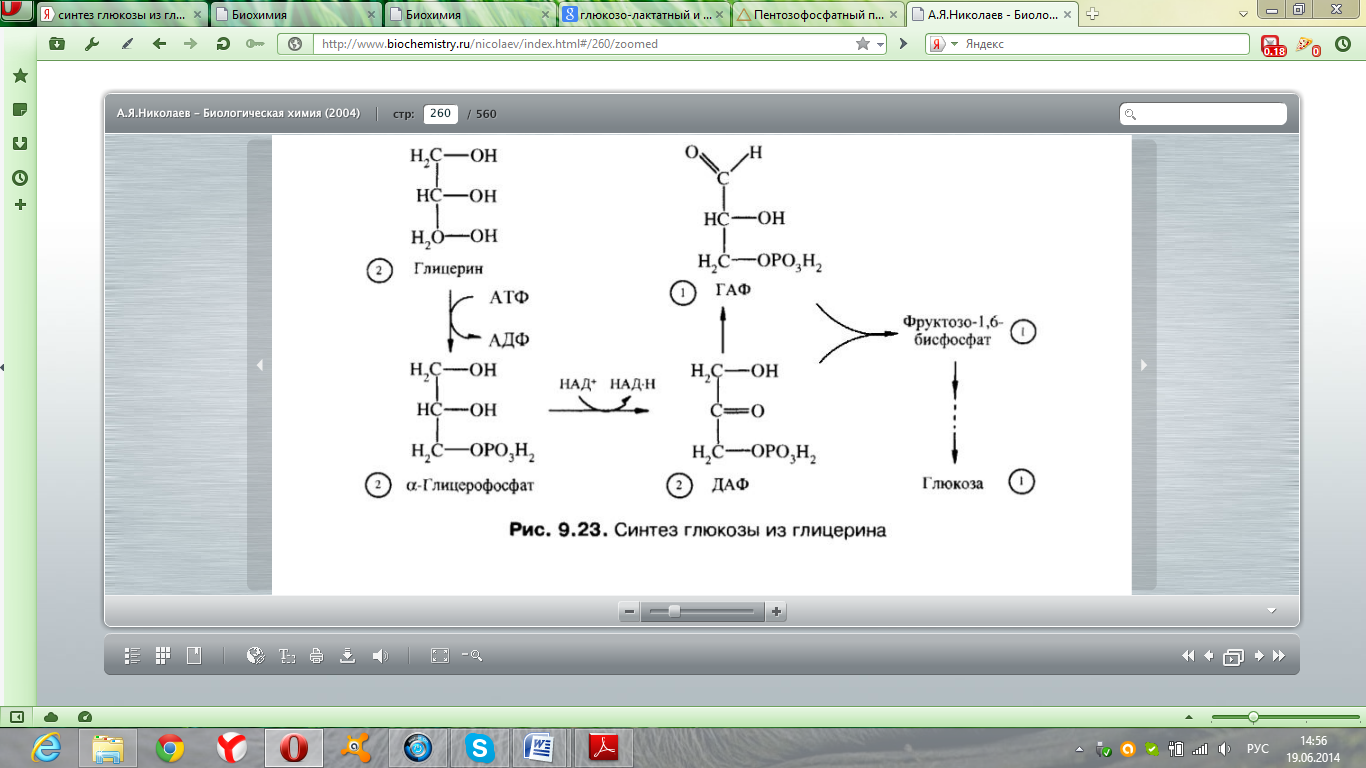 Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюкозонеогенез. Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах → пируват в мышцах → аланин в мышцах → аланин в печени → глюкоза в печени → глюкоза в мышцах. Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз. Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюкозонеогенез. Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах → пируват в мышцах → аланин в мышцах → аланин в печени → глюкоза в печени → глюкоза в мышцах. Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз.Глицерол образуется при гидролизе триацил-глицеролов, главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицерол киназа, например печень, почки. Этот АТФ-зависимый фермент катализирует превращение глицерола в α-глицерофосфат (глицерол-3-фосфат). При включении глицерол-3-фосфата в глюконеогенез происходит его дегидрирование NAD-зависимой дегидрогеназой с образованием дигидроксиацетонфосфата, который далее превращается в глюкозу. Глюконеогенез (ГНГ) ГНГ – синтез глюкозы из неуглеводных предшественников
ГНГ необходим для поддержания нормальной концентрации глюкозы в крови (для работы мозга) при длительном голодании и длительных физических нагрузках. Локализация – в цитоплазме и частично митохондриях клеток печени; немного в надпочечниках (не имеет энергетического значения) Сущность: обратный гликолизу процесс (ПВК → глюкоза) за исключение 1, 3,10 необратимых реакций. Эти реакции будут протекать по другому (обходные реакции). Регуляторный фермент, аллостерический, определяющий скорость реакции – фруктозо-1,6-бисфосфатаза. Из 2 ПВК для образования 1 глюкозы нужно 6 АТФ (4 АТФ и 2 ГТФ). Активируется кортизолом (гормон надпочечников) и зависит от витамина Н (биотина). 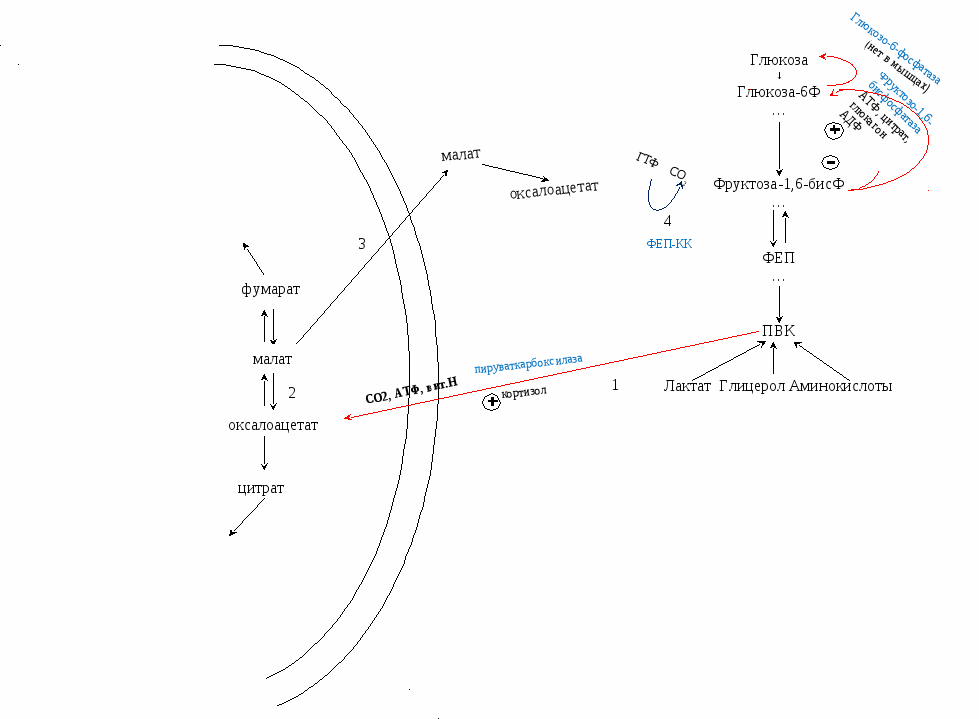  Первая обходная реакция:
ПВК из цитоплазмы поступает в митохондрию. Там он превращается в оксалоацетат (ЦТК) под действием фермента пируваткарбоксилазы (VI класс) АТФ, СО2 и витамина Н. Фермент активируется кортизолом.
Далее реакции ГНГ идут в обратном направлении гликолизу. Вторая обходная реакция: Фруктоза-1,6-бисФ превращается в фруктозу-6Ф под действием фермента фруктоза-1,6-бисфосфатаза (регуляторный, аллостерический). Третья обходная реакция: Глюкоза-6Ф превращается в глюкозу под действием фермента глюкоза-6-фосфатаза. |