УЧЕНИЕ О НЕРВНОЙ СИСТЕМЕ. Учение о нервной системе неврология общая неврология
 Скачать 11.49 Mb. Скачать 11.49 Mb.
|

|
Рис. 245. Спинно-мозжечковые пути: а - задний: 1 - поперечный разрез спинного мозга; 2 - поперечный разрез продолговатого мозга; 3 - кора мозжечка; 4 - рубчатое ядро; 5 - шаровидное ядро; 6 - синапс в коре червя мозжечка; 7 - нижняя мозжечковая ножка; 8 - дорсальный (задний) спинно-мозжечковый путь; 9 - чувствительный узел спинномозгового нерва; б - передний: 1 - поперечный разрез спинного мозга; 2 - передний спинномозжечковый путь; 3 - поперечный разрез продолговатого мозга; 4 - синапс в коре червя мозжечка; 5 - шаровидное ядро; 6 - кора мозжечка; 7 - зубчатое ядро; 8 - спинномозговой узел 3. Проводящие пути болевой и температурной чувствительности от кожи шеи, туловища и конечностей организованы в соответствии с принципом сегментарности. Эти пути составляют трехнейронную цепь (рис. 246). Тела первых - рецепторных нейронов расположены в чувствительных узлах спинномозговых нервов и представлены псевдоуниполярными клетками. Периферические отростки первых нейронов контактируют с рецепторными комплексами. Центральные отростки в составе задних корешков входят в спинной мозг и синаптически оканчиваются на клетках собственного ядра, располагающихся в пределах заднего рога и являющихся вторыми нейронами тракта. Далее аксоны вторых нейронов в восходящем направлении переходят на противоположную сторону спинного мозга, формируя латеральный спиноталамический путь. В продолговатом мозге латеральный спиноталамический путь, присоединяясь к нервным волокнам медиальной петли, достигает тела третьего нейрона, которое расположено в таламусе промежуточного мозга. В нейронах таламуса осуществляются сложная многопрофильная обработка поступившей от вторых нейронов информации и ее адаптация для следующего уровня. Аксоны третьих нейронов проходят через внутреннюю капсулу и в составе нервных волокон лучистого венца достигают зоны центрального отдела кожного анализатора в постцентральной извилине коры конечного мозга.
4. Проводящие пути болевой и температурной чувствительности от головы располагаются на основе тройничного нерва. В составе пути имеются 3 уровня, обозначенные тремя нейронами. Тела рецепторных - первых 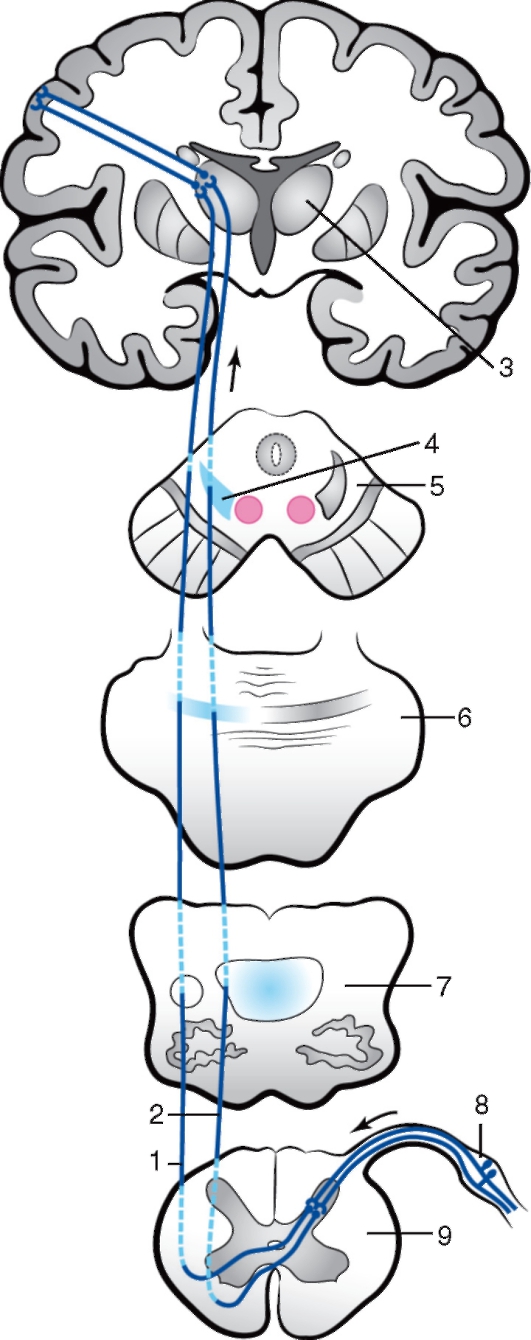 Рис. 246. Проводящие пути болевой и температурной чувствительности, осязания и давления (схема): 1 - латеральный спиноталамический путь; 2 - передний спиноталамический путь; 3 - таламус; 4 - медиальная петля; 5 - поперечный разрез среднего мозга; 6 - поперечный разрез моста; 7 - поперечный разрез продолговатого мозга; 8 - спинномозговой узел; 9 - поперечный разрез спинного мозга. Стрелками показано направление движения нервных импульсов нейронов локализованы в тройничном узле. Дендриты, или периферические отростки первых нейронов, имеются в составе каждой из трех главных ветвей тройничного нерва. Центральные отростки первых нейронов в составе тройничного нерва входят в мост, а потом образуют нисходящий пучок нервных волокон, достигающих вторых нейронов, формирующих ядро спинномозгового пути тройничного нерва. Аксоны вторых нейронов, переходя на контралатеральную половину, формируют основу тройничной петли, располагающейся рядом с медиальной петлей и достигающей соответствующего ядра таламуса, образованного телами третьих нейронов. После анализа и интегративной обработки информации в пределах уровня третьего нейрона нервные импульсы распространяются по аксонам третьих нейронов во внутреннюю капсулу и лучистый венец к центральному отделу кожного анализатора в область постцентральной извилины. 5. Проводящие пути тактильной чувствительности (сознательной) являются трехнейронными. Рецепторные - первые - нейроны располагаются в чувствительных узлах спинномозговых нервов и представляют собой псевдоуниполярные нервные клетки. Каждый нейрон имеет периферический отросток - дендрит, контактирующий с рецепторами кожи, и центральный отросток - аксон, вступающий в составе задних корешков в спинной мозг.
Преобладающая часть аксонов первого нейрона оканчивается на нейронах студенистого вещества, расположенных в заднем роге спинного мозга и являющихся вторыми нейронами пути. Сравнительно небольшая часть вторых нейронов располагается в составе ядер тонкого и клиновидного пучков в продолговатом мозге. Чтобы достичь их, аксоны первых нейронов, минуя серое вещество спинного мозга, поднимаются вверх в составе задних канатиков. Аксоны вторых нейронов обеих частей пути достигают следующего уровня - таламуса (третий нейрон), проходя в составе афферентных пучков, сопровождающих медиальную петлю. Следует подчеркнуть, что большая часть аксонов вторых нейронов, локализованная в задних рогах спинного мозга, до вступления в продолговатый мозг переходит на восходящем сегментарном уровне на контралатеральную сторону и формирует передний спиноталамический тракт в составе боковых канатиков. Аксоны третьих нейронов достигают центрального отдела анализатора в области постцентральной извилины, проходя в составе внутренней капсулы через лучистый венец к коре конечного мозга. 6. Проводящие пути стереогноза, или пространственной кожной чувствительности, участвуют в обеспечении узнавания предметов ощупыванием и являются трехнейронными. Тела рецепторных, или первых, нейронов традиционно участвуют в образовании чувствительных узлов спинномозговых нервов и представлены псевдоуниполярными клетками. Их периферические отростки в составе производных спинномозговых нервов контактируют с рецепторными зонами. Аксоны первых нейронов входят в спинной мозг как часть задних корешков и, не переключаясь на нейронах задних рогов, вступают в состав тонкого и клиновидного пучков задних канатиков. В составе ядер тонкого и клиновидного пучков в продолговатом мозге располагаются тела вторых нейроновпроводящего пути стереогноза.
В свою очередь аксоны вторых нейронов присоединяются к нервным волокнам медиальной петли и в ее составе переходят на противоположную сторону. Следующий уровень проводящих путей пространственной кожной чувствительности, или третий нейрон, располагается в таламусе промежуточного мозга. Отсюда аксоны третьих нейронов в составе внутренней капсулы, а затем лучистого венца достигают центрального отдела анализатора в коре конечного мозга (верхней теменной дольки). 7. Обонятельные проводящие пути обеспечивают восприятие запахов. Первыми нейронами являются особые нейросенсорные клетки обонятельной области полости носа. Их аксоны через решетчатую кость проникают из полости носа в полость черепа, где контактируют совторыми нейронами в обонятельной луковице. Аксоны вторых нейронов образуют обонятельных тракт, подходящий к телам третьих нейронов в обонятельном треугольнике. Оттуда волокна идут в кору - обонятельный мозг. 8. Проводящие пути от органов растительной жизни - внутренних органов, кровеносных и лимфатических сосудов и др. - образуют цепи из 3 нейронов. Первыми нейронами этих путей являются клетки узлов некоторых черепных нервов (тройничного, лицевого, языкоглоточного и блуждающего) и чувствительных узлов спинномозговых нервов, вторые нейроны - это клетки ядер указанных нервов и задних, а также боковых рогов спинного мозга; третьи - клетки таламуса, отростки которых заканчиваются в нижней части постцентральной извилины, в лобной доле впереди предцентральной извилины, лимбических образованиях, а также в других участках коры полушарий. В спинном мозге и стволе мозга данные проводящие пути проходят в определенной степени диффузно, без образования компактных самостоятельных трактов. В связи с перечисленными особенностями обеспечивается устойчивость интероцептивного анализатора, выполняющего функции поддержания жизнедеятельности, к локальным повреждениям.
Эфферентные проводящие пути 1. Корково-спинномозговой (пирамидный) и корково-ядерный пути (рис. 247) осуществляют эфферентную иннервацию произвольной поперечнополосатой (скелетной) мускулатуры. Оба пути состоят из двух нейронов: центрального и периферического. Центральный нейрон находится в двигательной области коры - предцентральной извилине, в пятом слое, и представлен гигантскими пирамиднымиклетками. При этом корково-ядерный путь начинается от нижней части данной извилины, а корково-спинномозговой - от верхней (см. рис. 234). Аксоны центральных нейронов проходят: корково-ядерного пути - в колене внутренней капсулы, а корково-спинномозгового - в передней части задней ножки капсулы. На протяжении мозгового ствола они располагаются преимущественно вентрально (спереди) по отношению к другим проводящим путям и заканчиваются на периферических нейронах. Тела периферических нейронов находятся у корково-ядерного пути в двигательных ядрах черепных нервов в стволе мозга. Двигательные ядра имеют III, IV, V, VI, VII, IX, X, XI и XII пары черепных нервов. Аксоны нейронов образуют указанные нервы и иннервируют мускулатуру головы, частично - шеи, а также глотки, мягкого нёба и языка. Вторыми нейронами корково-спинномозгового пути являются клетки передних рогов спинного мозга, аксоны которых в составе спинномозговых нервов иннервируют мышцы туловища, конечностей и частично шеи. Необходимо отметить, что аксоны первого нейрона корково-ядерного пути идут ко вторым нейронам не своей, а противоположной стороны. Большая часть волокон корково-спинномозгового пути совершает перекрест в нижнем отделе продолговатого мозга (decussatio pyramidum) и переходит в боковой канатик спинного мозга, где образует латеральный (пирамидный) корково-спинномозговой путь (tr. corticospinalis lateralis).Волокна этого пути заканчиваются в двигательных ядрах передних рогов спинного мозга. Меньшая часть волокон в области продолговатого мозга перекреста не образует. Эти волокна в спинном мозге составляет передний корково-спинномозговой путь
(tr. corticospinalis ventralis), часть волокон которого заканчивается в двигательных ядрах спинного мозга своей стороны, а другая часть волокон через переднюю белую спайку совершает перекрест и заканчивается в таких же ядрах противоположной стороны. Передний пучок прослеживается только в шейных и верхних грудных сегментах спинного мозга. Таким образом, пирамидный путь осуществляет преимущест- 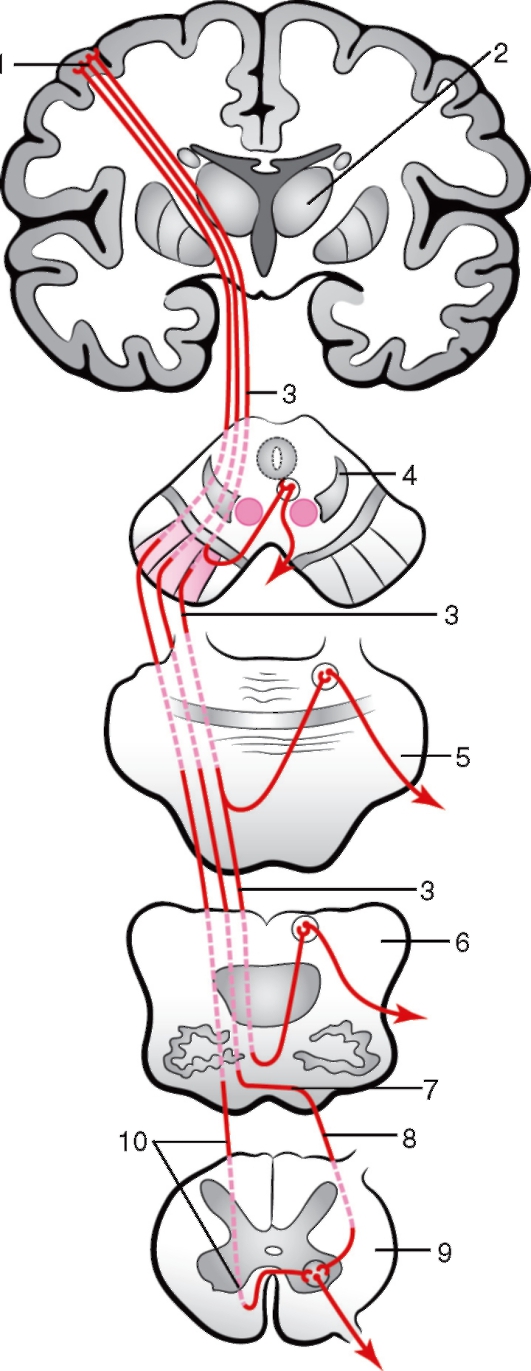 Рис. 247. Схема пирамидных проводящих путей: 1 - предцентральная извилина; 2 - таламус; 3 - корково-ядерный путь; 4 - поперечный разрез среднего мозга; 5 - поперечный разрез моста; 6 - поперечный разрез продолговатого мозга; 7 - перекрест пирамид; 8 - латеральный корково-спинномозговой путь; 9 - поперечный разрез спинного мозга; 10 - передний корково-спинномозговой путь. Стрелками показано направление движения нервных импульсов венно перекрестную иннервацию мускулатуры тела, и при одностороннем поражении центральных нейронов любого отдела наступает паралич мышц на противоположной очагу поражения стороне. Такая односторонняя иннервация характерна не для всех мышечных групп. Большая часть мышц, а именно: мышцы глазного яблока, жевательные, мышцы верхней части лица, глотки, гортани, шеи, туловища и промежности, получают так называемую двустороннюю иннервацию, т.е. от центральных нейронов как своей, так и противоположной стороны. Это в известной степени обеспечивает сохранение иннервации этих мышц при одностороннем поражении центральных нейронов. Односторонняя иннервация - у мышц конечностей, языка и мимических мышц нижней половины лица. В связи с этим при одностороннем поражении центральных нейронов этих мышц полностью утрачивается способность к движению, т.е. наступает паралич. 2. Проводящие пути экстрапирамидной системы проходят к передним рогам спинного мозга вне пирамидных путей, через покрышку и ретикулярную формацию мозгового ствола. К экстрапирамидной системе относят хвостатое и чечевицеобразное ядра, таламус, черное вещество, красное ядро, образования гипоталамуса, мозжечок, нижнюю оливу и ядра ретикулярной формации мозгового ствола. Экстрапирамидная система является филогенетически более старым образованием по сравнению с пирамидной. Функция экстрапирамидной системы заключается в осуществлении сложных безусловнорефлекторных двигательных актов, основанных на опыте вида, а двигательная сознательная корковая деятельность основана на условных рефлексах, закрепляющих индивидуальный опыт.
Экстрапирамидная система обеспечивает автоматические, без участия сознания, регуляцию и координацию тонуса мышечной ткани, осуществляет сокращение или расслабление определенных групп мышц, обеспечивает готовность двигательного аппарата к выполнению сложных, точных, целенаправленных действий. Поражения экстрапирамидной системы проявляются главным образом изменениями мышечного тонуса и двигательной активности. Основными проводящими путями экстрапирамидной системы являются: красноядерно-спинномозговой (рис. 248), покрышечно-спинномозговой, ретикулоспинномозговой, преддверно-спинномозговой, идущие от соответствующих образований к мотонейронам передних рогов спинного мозга. 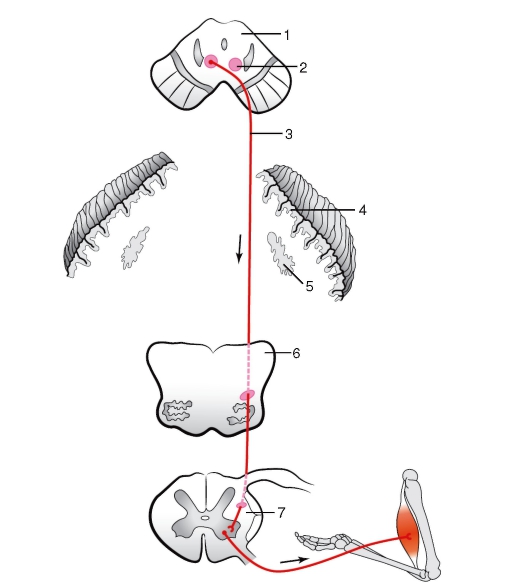 Рис. 248. Красноядерно-спинномозговой путь (схема): 1 - разрез среднего мозга; 2 - красное ядро; 3 - красноядерно-спинномозговой путь; 4 - кора мозжечка; 5 - зубчатое ядро; 6 - разрез продолговатого мозга; 7 - разрез спинного мозга. Стрелками показано направление движения нервных импульсов 3. Проводящие пути, регулирующие деятельность органов растительной жизни, обеспечивают функционирование внутренних органов и сосудов. Общая координация осуществляется корой полушарий (нижняя часть постцентральной извилины, лобная доля впереди предцентральной извилины, лимбическая система и некоторые другие участки). Оттуда импульсы поступают в подкорковые центры: базальные ядра, а также мозжечок. Основным образованием, специализирующимся на выполнении указанных функций, является гипоталамус. Третьим, более низким уровнем в пределах ЦНС, являются ядра ствола головного и спинного мозга. К ним относятся автономные (вегетативные ядра черепных нервов - III, VII, IX, X пары), ядра боковых рогов спинного мозга, а также ретикулярная формация. Перечисленные выше образования связываются между собой ассоциативными и проекционными нервными волокнами.
Типовые особенности строения центральной нервной системы Центральная нервная система у разных людей имеет некоторые специфические структурно-функциональные различия. Эти различия касаются формы мозга и его отдельных образований, величины, массы и других показателей. Так, масса головного мозга человека, как правило, составляет от 1100 до 2300 г. Следует отметить, что большая масса мозга (в пределах нормы) не свидетельствует о высоком интеллекте, и наоборот. Между формой головного мозга и формой черепа наблюдается определенное соответствие. Долихоцефалической, мезоцефалической и брахицефалической формам черепа соответствует определенное положение его отделов, образований, борозд и т.д. Это обстоятельство имеет важное практическое значение в связи с возможностью проекций на наружные покровы головы различных ядер, борозд и извилин. Оболочки головного мозга Твердая оболочка головного мозга (dura materencephali) прилегает к внутренней поверхности костей черепа и плотно сращена с ней в области основания и швов. Рыхлое соединение оболочки с костями свода черепа послужило основанием для выделения очень узкого эпидурального пространства, которое пронизано соединительнотканными волокнами, сосудами и нервами. К гладкой внутренней поверхности твердой оболочки прилегает паутинная оболочка головного мозга. Узкую щель между ними условно называют субдуральным пространством (рис. 249). В некоторых местах твердая оболочка образует отроги (отростки), которые проникают между различными отделами головного мозга. Так, в продольной щели, разделяющей полушария, расположен серповидной формы отросток - серп большого мозга (falx cerebri). 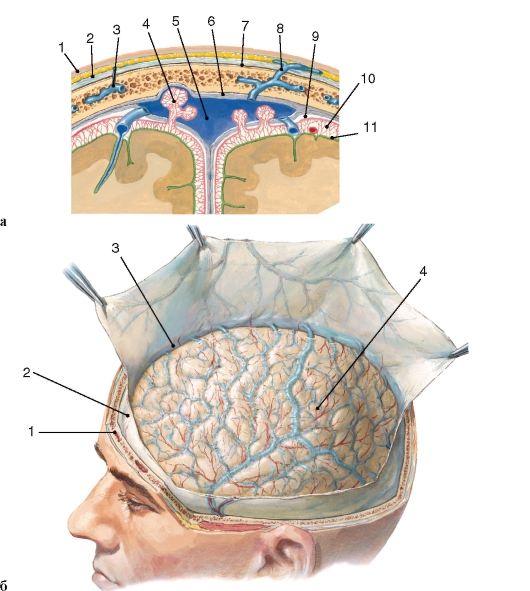 Рис. 249. Оболочки мозга: а - срез через структуры головы в области свода черепа: 1 - кожа; 2 - сухожильный шлем; 3 - диплоическая вена; 4 - грануляция паутинной оболочки; 5 - верхний сагиттальный синус; 6 - твердая мозговая оболочка; 7 - эпидуральное пространство; 8 - эмиссарная вена; 9 - паутинная оболочка; 10 - подпаутинное пространство; 11 - мягкая оболочка;
б - вид оболочек мозга после вскрытия черепа: 1 - кость черепа; 2 - твердая оболочка; 3 - верхний сагиттальный синус; 4 - паутинная оболочка, через которую просвечивают артерии (красного цвета) и вены (синего), залегающие в подпаутинном пространстве на мягкой оболочке |