Нейротропные средства. Нейротропные средства (1). Введение определение
 Скачать 0.77 Mb. Скачать 0.77 Mb.
|

“Мишени” для нейротропных веществНа пресинаптическом уровне различные биологически- активные вещества могут влиять на процессы синтеза, де- понирования и выделения медиатора, усиливая или тормозя эти процессы. Вещества могут прямо взаимодействовать с рецепторными структурами, в результате чего они или имитируют эффек- ты медиатора или препятствуют взаимодействию медиато- ра с рецептором, т.е. могут вести себя как агонисты или антагонисты. С учетом структуры и функционирования синапса можно предположить следующие точки, воздействуя на которые можно модулировать синаптическую передачу: 3. Точкой приложения для некоторых фармакологических веществ на уровне синапса, могут быть системы инактива- ции медиатора. При этом вещества ослабляющие эти си- стемы, как правило, усиливают передачу нервного возбуждения в синапсе, а вещества повышающие актив- ность процессов инактивции - ослабляют передачу в синап- се. Направленность действия вещества или знак синаптического действия может зависеть не только от точки приложения или характера взаимодействия с той или иной структурой, но и от концентрации вещества и времени, т.е. может иметь место временная фазность в действии вещества. Глава 2ХОЛИНЕРГИЧЕСКИЙ СИНАПСМедиатор (Ацетилхолин). Ацетилхолин - медиатор нервных импульсов в различных си- напсах НС. Синтез ацетилхолина происходит в аксоплазме холинергических нервов из активной уксусной кислоты (аце- тилкоэнзим-А) и аминоспирта холина и катализируется фер- ментом холинацетилазой. 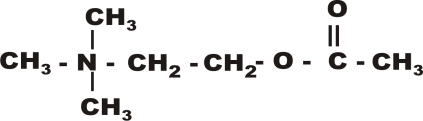 Ацетилхолин депонируется в связанной с белком форме и накапливается в непосредственной близости от пресинаптиче- ской мембраны (Рисунок 3). Выброс АХ происходит как под действием приходящего нервного импульса, так и спонтанно (по крайней мере, в нерв- но-мышечном синапсе) в виде квантов, содержащих несколь- ко тысяч молекул медиатора. Спонтанное выделение медиатор занимает важное место в поддержании функциональной го- товности мышцы. Существенную роль в процессе выделения медиатора играет деполяризация пресинаптической мембраны и вхождение в клетку ионов Са++. Помимо собственно медиаторной функции, АХ играет роль гуморального фактора, участвуя в реализации некоторых вне- синаптических эффектов. Так, он вызывает освобождение ионов калия, связанных с белками, изменяет проницаемость биомембран, участвует в регуляции селективной проницаемо- сти эритроцитов, регулирует активность некоторых дыхательных ферментов, влияет на метаболизм макроэргических фосфатных соединений и т.д. Вероятно, в основе многих немедиаторных эффектов АХ лежит его спо- собностью изменять проницаемость мембран для ионов, осо- бенно Са++.  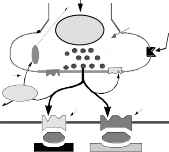  Х о л и н а ц е т и л а з а х о л и н + а ц е т и л - К о А Х о л и н а ц е т и л а з а х о л и н + а ц е т и л - К о АА Х + б е л о к Са ++ а д р е н а л и н Х о л и н А Х Э Н- Х р( + ) АХ А Х АХ М - Х р ( -) - А д Р ( + ) А ц е т а т н М - Х Р Н - Х Р Рисунок 3. Строение холинергического синапса АХ - ацетилхолин; АХЭ - ацетилхолинэстераза; ХР - хо- линорецептор. ХолинорецепторПод термином холинорецептор (ХР) в дальнейшем мы будем подразумевать белковую макромолекулу, способную избира- тельно взаимодействовать АХ, причем это взаимодействие вызывает краткотекущее изменение физико-химических свойств мембраны, в которую ХР встроен. Структура холинорецептора В литературе наиболее часто приводится строение холиноре- цептора по Е. Д. ДеРобертису (1976), который представил его в виде тетрамера, расположенного на мембране и связанного с фосфатидилинозитидом. Последний образует канал-ионофор, открытие которого происходит при изменении четвертичной структуры рецептора. Существуют и некоторые другие, но близкие, представления о структуре холинорецептора, например, по Кистлеру и Шпроу- ду в учебнике под ред. Харкевича.  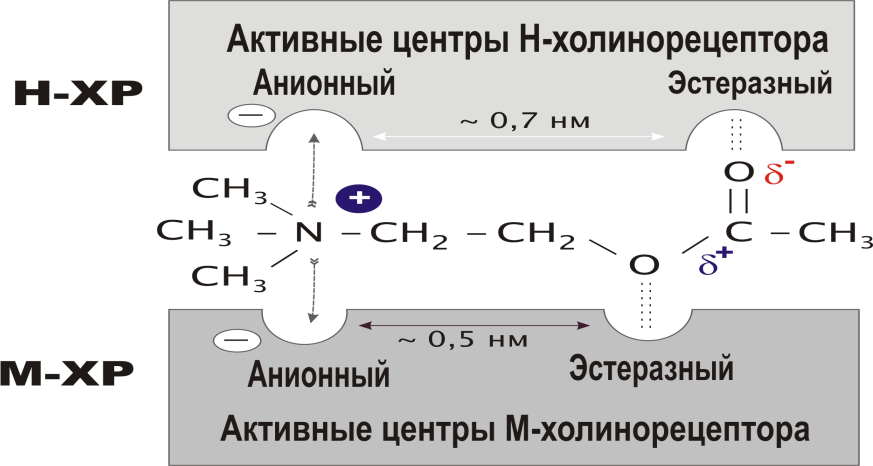 Рисунок 4. Схема взаимодействия ацетилхолина с детекторной субъеденицей холинорецептора (по Кушински и Люльман). Описа- ние в тексте. Холинорецептор имеет два активных центра: отрицательно заряженный анионный центр, в который вхо- дят карбоксильные группы глутамата или аспаргината; этот центр вступает во взаимодействие с положительной кати- онной головкой АХ; эстерофильный центр, взаимодействующий с кислородом карбонильной группой или кислородом эфирной связи. Молекула ацетилхолина, в свою очередь, содержит 3 про- странственно разобщенных центра, имеющих значение для его биологической активности. Ацетилхолин взаимодействует с холинорецептором всегда с помощью двух центров, одним из которых обязательно является катионная головка. Вторым центром взаимодействия может быть «карбонильный» или «эфирный» кислород. В этом, по-видимому, состоит отличие между М- и Н-холинорецепторами. Предполагают тесную связь активации М-холинорецепторов с системой фосфотидилинозитола и гуанилатциклазы и, как следствие этого повышение концентрации клеточного цГМФ, который, как известно, является вторичным “мессенджером” в клетке. Это может быть следствием и того, что открываются связанные с рецептором Са++-инофоры (потенциалнезависи- мые каналы) и увеличивается внутриклеточная концентрация Са++. В настоящее время идентифицированы и так называемые пре- синаптические рецепторы, располагающиеся на преси- наптических окончаниях нервов. Согласно гипотезе T. Полак (1971), пресинаптические М-холинорецепторы регулируют освобождение АХ по принципу отрицательной обратной свя- зи, т.е. их стимуляция снижает высвобождение медиатора. Роль пресинаптических Н-рецепторов, обнаруженных в нерв- но-мышечном аппарате и ЦНС обратная, их стимуляция уси- ливает секрецию АХ. На пресинаптическом окончании холинергических волокон обнаружены рецепторы и к другим химическим модуляторам. Существуют и внесинаптические холинорецепторы. В конце 70-х годов появились данные о наличии холинорецепторов на иммунокомпетентных клетках и эритроцитах. Инактивация ацетилхолина Образовавшийся при взаимодействии комплекс АХ-ХР бы- стро распадается и выделившийся АХ подвергается фермен- тативному гидролизу ацетилхолинэстеразой (АХЭ). Разру- шение АХ происходит или в синаптической щели, или на постсинаптической мембране. Существуют два вида холине- стеразы: ацетилхолинэстераза (истинная ХЭ) и бутирилхо- линэстераза (ложная ХЭ). АХЭ располагается внутрисинап- тически, ложная в плазме и экстрацеллюлярной жидкости. Механизм гидролиза АХ следующий. Молекула ацетилхолина является полярной: триметиламмониевая группировка (кати- онная головка) имеет положительный заряд, а карбоксильная - отрицательный. В активном центре ХЭ также два участка, не- сущих заряд: анионный - образованный аспаргиновой кисло- той, и эстеразный пункт, в который входят серин, гистидин и тирозин. Катионная головка молекулы ацетилхолина фикси- руется на анионном пункте активного центра холинэстеразы, а эстеразный пункт ХЭ образует ковалентную связь с углеродом карбоксильной группы АХ. Происходит разрыв эфирной свя- зи в молекуле АХ с образованием свободного холина (частич- но захватывается пресинаптическим образованием) и ацетилированного фермента, который очень быстро регенери- рует (Рисунок 3). Существует достаточно обоснованное мнение, что наряду с ферментативным разрушением АХ, в некоторых холинергиче- ских синапсах (ганглии?) присутствует и механизм инактива- ции по типу экстранеронального захвата или удаления АХ из синаптического пространства. Однако следует признать, что роль его невелика. |