Зачем врачу нужна биологическая химия
 Скачать 6.47 Mb. Скачать 6.47 Mb.
|

Длительная стабилизация сдвигов рНФизиологическая компенсация нарушений КОСДыхательная системаЛегочная вентиляция обеспечивает удаление угольной кислоты, образованной при функционировании бикарбонатной буферной системы. По скорости реакции на изменение рН – это вторая система после буферных систем. Дополнительная вентиляция легких приводит к удалению СО2, а значит и Н2СО3, и повышает рН крови, что компенсирует закисление межклеточной жидкости и плазмы крови продуктами метаболизма, в первую очередь, органическими кислотами. Сдвиги значений рО2 не являются сильно значимыми для изменения легочной вентиляции. Только снижение рО2 до 8 кПа в артериальной крови (норма 11,04-14,36 кПа или 83-108 мм рт.ст.) приводит к увеличению активности дыхательного центра. Более существенным фактором для активации дыхательной системы является концентрация ионов Н+. Накопление ионов Н+ в крови уже через 1-2 минуты вызывает максимальную (для данной их концентрации) стимуляцию дыхательного центра, повышая его активность до 4-5 раз, что приводит к снижению рСО2 до 10-15 мм рт.ст. И, наоборот, снижение кислотности крови понижает активность дыхательного центра на 50-75%, рСО2 при этом способен возрастать до 60 мм рт.ст и выше. Костная тканьЭто наиболее медленнореагирующая система. Механизм ее участия в регуляции рН крови состоит в возможности обмениваться с плазмой крови ионами Са2+ и Na+ в обмен на протоны Н+. Происходит растворение гидроксиапатитных кальциевых солей костного матрикса, освобождение ионов Са2+ и связывание ионов НРО42– с Н+ с образованием дигидрофосфата, который уходит с мочой. 8Н+ + Ca10(PO4)6(OH)2 → 2H2O + 10Ca2+ + 6HPO42– Параллельно при снижении рН (закисление) происходит поступление ионов H+внутрь остеоцитов, а ионов калия – наружу. ПеченьСущественную, но пассивную роль в регуляции кислотно-основного состояния крови берет на себя печень, в которой происходит метаболизм низкомолекулярных органических кислот (молочная кислота и др). Кроме этого, кислые и щелочные эквиваленты выделяются с желчью. ПочкиРазвитие почечной реакции на смещение кислотно-основного состояния происходит в течение нескольких часов и даже дней. Специфические нейрогуморальные механизмы регуляции секреции и реабсорбции ионов Н+отсутствуют. Регуляция концентрации ионов H+ осуществляется опосредованно, через потоки ионов Na+, движущихся по градиенту концентрации, и через перераспределение потоков ионов К+ и Н+, которые выходят из эпителиоцитов (секретируются) в обмен на ионы Na+. Также для обеспечения электронейтральности внутри- и внеклеточной жидкости при реабсорбции ионов Na+усиливается реабсорбция ионов Cl–, однако их не хватает, поэтому возникает необходимость в усилении реабсорбции и дополнительном синтезе ионов HCO3–. Все вышесказанное упрощенно можно проиллюстрировать выражением: Реабсорбция (Na+) ≈ Секреция (К+ + Н+) + Реабсорбция (Cl– + HCO3–) В почках активно протекают три процесса, связанных с уборкой кислых эквивалентов. Благодаря этим процессам рН мочи в состоянии снижаться до 4,5-5,2 :
Реабсорбция бикарбонат-ионовВ проксимальныхканальцах ионы Na+ мигрируют в цитозоль эпителиальных клеток в силу концентрационного градиента, который создается на базолатеральной мембране при работе фермента Na+,К+ АТФазы. В обмен на ионы Na+ эпителиоциты канальцев активно секретируют в канальцевую жидкость ионы водорода. Ионы HCO3– первичной мочи и секретируемые ионы Н+ образуют угольную кислоту Н2СО3. В гликокаликсеэпителиоцитов фермент карбоангидразакатализирует распад угольной кислоты на СО2 и воду. В результате возникает градиент концентрации углекислого газа между просветом канальцев и цитозолем и СО2диффундирует в клетки. Внутриклеточнаякарбоангидраза использует пришедший СО2 и образует угольную кислоту, которая диссоциирует. Ионы НСО3– транспортируются в кровь, ионы Н+ – секретируются в мочу в обмен на ионы Na+. Таким образом, объем реабсорбции НСО3– полностью соответствует секреции ионов Н+. 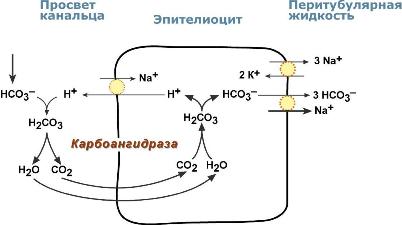 Процесс реабсорбции бикарбонат-ионовВ проксимальныхканальцах происходит реабсорбция 90% профильтрованного НСО3–. В петле Генле и дистальных канальцах реабсорбируется оставшееся количество карбонат-иона. Всего в почечных канальцах реабсорбируется более 99% от фильтруемых бикарбонатов. АцидогенезВ процессе ацидогенеза в сутки с мочой выделяется 10-30 ммоль кислот, называемых титруемыми кислотами. Фосфаты, являясь одной из этих кислот, играют роль буферной системы в моче. Роль этой системы состоит экскреции кислых эквивалентов без потерь бикарбонат-ионов за счет дополнительного иона водорода в составе выводимого NaH2PO4 (по сравнению с NaHCO3): Na2HPO4 + Н2СО3 → NaH2PO4 + NaНСО3 После того как бикарбонат натрия в почечных канальцах реабсорбируется, кислотность мочи зависит только от связывания ионов Н+ с HPO42– и содержания дигидрофосфата. 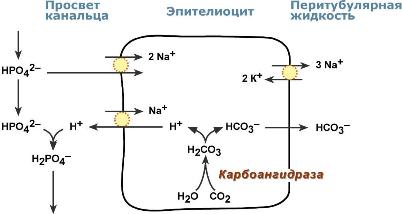 Процесс ацидогенеза в почечных канальцахХотя в крови соотношение HРO42– : H2РO4– равно 4 : 1, в клубочковом фильтрате оно меняется на 1 : 9. Происходит это из-за того, что менее заряженный H2РO4? лучше фильтруется в клубочках. Связывание ионов Н+ ионами HРO42– на протяжении всего канальца приводит к увеличению количества H2РO4–. В дистальных канальцах соотношение может достигать 1 : 50. АммониегенезАммониегенез происходит на протяжении всего почечного канальца, но более активно идет в дистальных отделах – дистальных канальцах и собирательных трубочках коркового и мозгового слоев. Глутамин и глутаминовая кислота, попадая в клетки канальцев, быстро дезаминируются ферментами глутаминазаиглутаматдегидрогеназа с образованием аммиака. Являясь гидрофобным соединением, аммиак диффундирует в просвет канальца и акцептирует ионы Н+ с образованием аммонийного иона. Источником ионов H+ первичной мочи в проксимальныхотделах канальца является Na+,H+-антипорт. Вдистальныхотделах, в отличие от проксимальных, секреция ионов Н+ происходит с участием Н+-АТФазы, локализованной на апикальной мембране вставочных клеток. 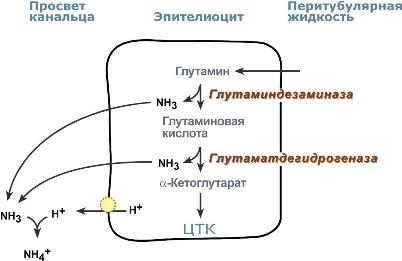 Процесс аммониегенеза в дистальных канальцахАммонийный катион NH4+ способен взаимодействовать с анионами Cl–, SO42–, с органическими кислотами (лактат и другие) с образованием аммонийных солей. |