|
|
Учебник по клиндиагностике (укр.). Учебник по клиндиагностике (укр. Вступ предмет клінічної діагностики
ВИЗНАЧЕННЯ РЕЗЕРВНОЇ ЛУЖНОСТІ І КИСЛОТНОЇ МІСТКОСТІ КРОВІ
Більшість біологічних процесів в організмі перебігає у нейтральному або слаболужному середовищі (за винятком травлення у шлунку), яке забезпечується рівновагою між кислотними (Н+) і лужними (ОН
) іонами і виражається водневим показником (рН). У плазмі крові величина рН постійна, утримується в межах 7,30—7,45.
На стан кислотно-основного балансу значно впливають як кислоти, що утворюються із білків і ліпідів у результаті проміжного обміну речовин у клітинах і тканинах, бродильних процесів в органах травлення, так і луги, які надходять в організм з рослинними кормами, утворюються в процесі обміну речовин (аміак,
262
аміни та ін.). Кислоти і луги також утворюються при розвитку різних патологічних процесів. Незважаючи на вплив кислот і лугів, водневий показник крові — величина досить стабільна. Постійність рН забезпечується насамперед буферними системами крові, а також активною діяльністю нирок, які виводять з організму лужні й кислі продукти обміну речовин, легень, що виділяють залишки вуглекислоти, органів травлення, шкіри.
У зв'язку з тим, що зрушення кислотно-лужного балансу в організмі компенсується, зміни концентрації водневих іонів спостерігаються лише в окремих, дуже тяжких випадках, і тому оцінку стану кислотно-лужної рівноваги провадять за результатами дослідження буферних систем крові: гідрокарбонатної, фосфатної, білкової та гемоглобінової. Найбільше значення мають бікарбонати і система гемоглобіну. В інтерстиціальній рідині, яка становить понад 2/з позаклітинної рідини організму, практично присутня лише гідрокарбонатна буферна система, а у внутрішньоклітинній рідині важливе значення має фосфатна.
Показниками кислотно-основного балансу є рН, парціальний тиск вуглекислоти (СО2), стандартний бікарбонат крові (СБ), буферні основи (БО) та їх зрушення (ЗБО), бікарбонат крові. Такі показники можна визначити в крові, взятій без доступу кисню, на приладах типу «Аструп», «Азів».
У практиці ветеринарної медицини такий аналіз провести важко, тому стан кислотно-основного балансу крові визначають за результатами дослідження резервної лужності плазми крові, одержаній без доступу повітря. Під резервною лужністю розуміють запас бікарбонатів крові, визначений за загальною кількістю вуглекислоти, яка міститься в основному в їх складі. Визначають резервну лужність плазми крові дифузним методом за допомогою здвоєних колб (за Кондрахіним І. П.). Величину її визначають кількістю мілілітрів вуглекислого газу, виділеного з 100 мл плазми крові й виражають в об'ємних процентах ССЬ. У здорових тварин резервна лужність становить 45—66 об% СО2.
Крім резервної лужності, в лабораторіях ветеринарної медицини визначають кислотну місткість сироватки або плазми крові за Беляєвим — Большаковим і Л. О. Кудрявцевою — В. Ф. Коро-мисловим. Кислотна місткість сироватки становить: у великої рогатої худоби — 230—380 мг/100 мл; коней — 270—400; овець — 230—320; свиней — 260—400; собак — 220—340 мг/100 мл.
При порушенні кислотно-основного балансу може розвиватися ацидоз (надлишок кислот і нестача лугів) або алкалоз (надлишок лугів). Якщо ацидоз і алкалоз перебігають без зрушення рН крові, то їх називають компенсованими, а якщо рН змінюється — декомпенсованими. За походженням ацидоз і алкалоз можуть бути метаболічними (обмінними) і респіраторними (газовими).
Метаболічний ацидоз зустрічається найчастіше. Він розвиває-
263
ться внаслідок порушень проміжного обміну речовин у тканинах і нагромадження в них органічних кислот (молочної, піровиноградної, ацетилоцтової та ін.) при недостатньому виділенні або розпаді цих метаболітів ураженими органами: печінкою, легенями, нирками, кишечником; при згодовуванні тваринам кормів, що містять надлишок органічних кислот, особливо масляної, при кон-центратному типі годівлі. У жуйних тварин причиною метаболічного ацидозу є згодовування великої кількості кормів, що містять надмірну кількість легкорозчинних вуглеводів — цукрових буряків, зернових концентратів, картоплі та інших, внаслідок чого утворюється значна кількість ЛЖК і молочної кислоти, розвивається ацидоз рубця.
Компенсований ацидоз розвивається також у випадку, коли тварини довго не користуються активним моціоном, внаслідок чого в організмі нагромаджуються недоокислені продукти обміну речовин, органічні кислоти, кетонові тіла, які зв'язують гідрокарбонати та інші лужні компоненти крові.
Метаболічний ацидоз є також наслідком різних захворювань — діареї різної етіології, рахіту, пневмонії, кетозу, хронічного ру-мініту, нефриту та нефрозу, серцево-судинної та дихальної недостатності, діабету й інших хвороб. Механізми розвитку метаболічного ацидозу бувають різними. Так, при діареях молодняка важливе значення мають виведення із організму великої кількості бікарбонатів, дегідратація, гіпоксія, порушення кровообігу, голодування. Внаслідок порушення постачання тканин киснем при захворюваннях легень і серця посилюються анаеробні процеси й утворення кислих продуктів, збільшується рівень кінцевих продуктів обміну речовин. При захворюваннях нирок метаболічний ацидоз зумовлюється зниженням рівня бікарбонатів і нагромадженням фосфатів, сульфатів і аніонів сильних органічних кислот у позаклітинній рідині, недостатньою секрецією іонів водню у сечу та іншими причинами.
Метаболічний ацидоз характеризується зниженням рН крові, парціального тиску вуглекислоти, бікарбонату і буферних основ крові. Значно виражений дефіцит буферних основ (ЗБО має негативний знак).
Метаболічний алкалоз характеризується нагромадженням в організмі основ або виведенням кислот з одночасним підвищенням рівня бікарбонатів у крові. Він буває наслідком порушення травлення в передшлунках, якщо тваринам згодовують надмірну кількість азотистих кормів (зеленої маси або сінажу конюшини, люцерни, протеїнових концкормів, сечовини і деяких солей амонію, особливо в поєднанні з недостатньою кількістю вуглеводистих кормів), або кормів, уражених плісеневими грибами, що призводить до відмирання симбіонтної мікрофлори, яка синтезує ЛЖК, і сприяє розмноженню гнильних бактерій. У великої рогатої худо-
264
би метаболічний алкалоз буває при зміщенні сичуга, а у моно-гастричних тварин — при блюванні, коли з масами втрачається соляна кислота.
Респіраторний ацидоз розвивається при недостатній вентиляції легень, коли не забезпечується повне виведення вуглекислоти із легень внаслідок підвищеного рівня рСОа в крові. В результаті цього в організмі збільшується вміст Н2СОз. Гіповентиляція легень буває при пневмонії, альвеолярній емфіземі, пневмотораксі, ателектазі легень, ексудативному плевриті, підвищенні внутрішньочеревного тиску при тимпанії рубця, гострому розширенні шлунка та метеоризмі кишечника, при ураженні дихального центру. Даний вид ацидозу супроводжується недостатнім надходженням кисню, що спричиняє гіпоксію, яка зумовлює також метаболічний ацидоз, тобто йде поєднання і респіраторного, й метаболічного ацидозу.
Респіраторний алкалоз характеризується дисбалансом між утворенням СО2 у тканинах і підвищеним виділенням його при гіпер-вентиляції легень. У хворих тварин вміст рСО2 в крові при цьому знижений. Гіпервентиляція легень виникає при безпосередньому впливу на дихальний центр, а також при енцефаліті, менінгіті, інтоксикації аміаком, пухлинах мозку, перегріванні організму, подразненні периферичних рецепторів в аорті і сонній артерії.
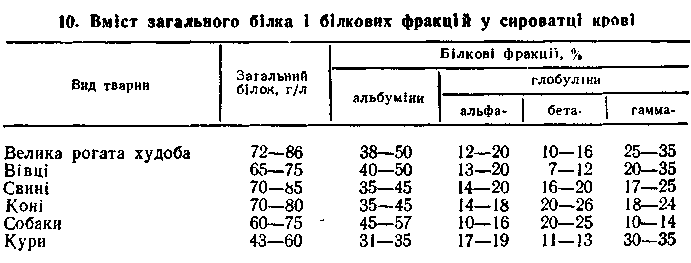
ВИЗНАЧЕННЯ ЗАГАЛЬНОГО БІЛКА І БІЛКОВИХ ФРАКЦІЙ
Білки сироватки крові — досить велика група білків, які різняться між собою структурою, фізико-хімічними властивостями, функціями. Загальну кількість їх визначають за допомогою рефрактометра або біуретовим методом, а окремі компоненти — електрофорезом. Залежно від методу розподілу білків можна одержати від 5 до 100 фракцій. Електрофорезом на папері в сироватці крові виявляють 4—5 фракцій: альбуміни, альфа- (інколи альфар і альфа2-), бета- і гамма-глобуліни, а електрофорезом в агаровому, крохмальному і поліакріламідному гелях — значно більше <до ЗО).
Кількість загального білка і співвідношення між окремими фракціями в сироватці крові тварин різних видів коливається у визначених межах (табл. 10).
У молодняка вміст загального білка нижчий, ніж у дорослих: у телят віком 1—10 днів — 56—70 г/л, новонароджених поросят — 45—50, ягнят — 46—54 г/л.
Зменшення вмісту загального білка в сироватці крові — гіпо-протеїнемія розвивається при недостатньому надходженні білків в організм, незбалансованості раціону за окремими незамінними амінокислотами, оскільки синтез білка обмежується тією аміно-
265
кислотою, якої надходить найменше (тому її називають лімітуючою); недостатньому перетравленні білка і всмоктуванні амінокислот у кишечнику, що зумовлене зниженням секреторної функції шлунка, кишечника, підшлункової залози, активності протеолітичних ферментів (при гастроентеритах, пневмоніях будь-якої етіології); порушенні синтезу білка в печінці при її хворобах (гепатит, гепатоз, цироз), оскільки в печінці синтезується 95— 100 % альбумінів; при втраті білка внаслідок захворювань нирок (нефроз, нефрит), кровотечі, обширних ексудатах і транссудатах, злоякісних пухлинах. Зменшення кількості білка в сироватці крові частіше всього відбувається за рахунок альбумінів — фракції, яка легко проходить через судинні мембрани та стінки клубочків нирок.
Гіперпротеїнемія зустрічається рідше. Вона може бути відносною (спостерігається при згущенні крові внаслідок втрати рідини при зневодненні) і абсолютною, яка майже завжди супроводжується збільшенням кількості гамма-глобулінів. Гіперпротеїнемія спостерігається при надмірному згодовуванні кормів, багатих на білки, гепатиті, хронічних інфекціях, токсикозах внаслідок подразнення імунокомпетентних клітин мононуклеарної системи, а також появи патологічних білків — парапротеїнів.
Крім вмісту загального білка, для діагностики різних патологічних процесів важливе значення має визначення білкових фракцій. Порушення оптимального співвідношення між ними називають диспротеїнемією. Найбільш виражені диспротеїнемії бувають у випадку ураження органів, де синтезуються білки. Особливо часто зменшується кількість альбумінів (гіпоальбу-мінемія), які виконують важливі функції щодо підтримання колоїдно-осмотичного тиску крові, регуляції водного обміну між кров'ю і міжтканинним простором, зв'язування та транспортування вуглеводів, ліпідів, гормонів, вітамінів, пігментів, мінеральних речовин.
Гіпоальбумінемія розвивається внаслідок білкового голодування і є типовою ознакою хвороб печінки (гепатит, гепатоз, абсце-
266
си, цироз і пухлини), оскільки в ній синтезуються альбуміни. Спостерігається вона при різних незаразних, інфекційних та паразитарних хворобах, коли настає вторинне ураження печінки (пневмонії, кетоз, перикардит, міокардоз, лейкоз, туберкульоз, сальмонельоз, колібактеріоз, диспепсія, гострі респіраторні хвороби та ін.). Особливо вираженою буває гігюальбумінемія при хронічних захворюваннях нирок (нефроз, нефрит), які супроводжуються втратою білка з сечею (протеїнурія) та розвитком набряків.
Збільшення кількості альбумінів буває рідко — в основному при дегідратації. При змінах кількості альбумінів порушується їх співвідношення з глобулінами (змінюється альбуміно-глобуліно-вий коефіцієнт), яке у здорових тварин коливається у межах від 0,7 до 1.
Кількість альфа-глобулінів збільшується при гострих запальних процесах (ревматизм, пневмонія, нефрит, артрит) та при загостренні хронічно перебігаючих хвороб (туберкульоз, гепатит), оскільки в цю групу входять білки «гострої фази» (С-реактивний білок, церулоплазмін, гаптоглобін, альфаі-антитрипсин, альфа2-макроглобулін, кислий альфаі-глікопротеїд), а зменшується їх рівень рідко, частіше всього при тяжких дистрофічних процесах у печінці, де частково синтезується альфа-глобулін.
Збільшення кількості бета-глобулінів спостерігається частіше при хронічноперебігаючих інфекціях, хворобах нирок (нефроз, нефрит), цирозі печінки. До складу фракцій бета-глобулінів входить фібриноген, збільшення вмісту якого буває при крупозній пневмонії, лейкозі, септичному ендокардиті, а зменшення — при хворобах печінки, де він синтезується.
Фракції гамма-глобулінів містять основну масу антитіл (імуно-глобулінів), які забезпечують гуморальний захист організму, тому кількість їх в сироватці крові залежить від морфологічної зрілості і функціональної повноцінності імунореактивної тканини.
Низький рівень гамма-глобулінів буває у новонароджених, особливо в перший день життя, оскільки гамма-глобуліни не проходять через плаценту, а надходять лише з молозивом (фізіологічний імунодефіцит), тому в підтриманні їх рівня має велике значення якість молозива, своєчасність його випоювання, стан слизової оболонки тонкого кишечника. Синтез власних імуноглобулі-нів починається з 5—7-го дня життя і досягає оптимального рівня лише в б-місячному віці, тому молодняк сприйнятливий до багатьох хвороб (сальмонельозу, стрептококозу, пастерельозу, гострих респіраторних, пневмоній). Зниження вмісту гамма-глобулінів спостерігається також при різних захворюваннях, які супроводжуються ураженням імунної системи (мієлома, лімфолейкоз, хвороба Гамборо), втратою імуноглобулінів при нефрозах, ентеритах, хронічних кровотечах, внаслідок пригнічення функції імунної системи різними токсинами, лікарськими препаратами (імунодопресанта-
267
ми), радіонуклідами, низьким рівнем у раціоні протеїну, біологічно активних речовин. Це так званий вторинний або набутий іму-нодефіцитний стан або вторинна гіпогаммаглобулінемія.
Гіпергаммаглобулінемія спостерігається при всіх імунологічних реакціях, які супроводжуються посиленим синтезом глобулінів (вакцинації), і зумовлена підвищенням вмісту майже всіх класів імуноглобулінів та неспецифічних антитіл, при багатьох бактеріальних інфекціях (стрепто-стафіло- і пневмококових), хронічних гепатитах, цирозі печінки, деяких паразитарних хворобах.
ВИЗНАЧЕННЯ КАРОТИНУ ТА ВІТАМІНУ А В СИРОВАТЦІ КРОВІ
Каротин е попередником вітаміну А. Він міститься в рослинних кормах, молозиві, молоці, жовтках яєць, печінці, риб'ячому жирі. Є кілька ізомерів каротину, серед яких найбільшу біологічну активність має бета-каротин. У стінці тонкого кишечника і частково у печінці каротин перетворюється у ретинол (вітамін А). Основним депо вітаміну А та каротину є печінка, де зберігається до 90 % наявного в організмі ретинолу.
Фізіологічні функції вітаміну А досить різнобічні. Всюди, де е орган зору, яким би примітивним він не був, вітамін А здійснює велику таємницю природи — перетворює фізичну енергію світла в зоровий нервовий сигнал. Вітамін А бере участь у формуванні і підтриманні структури епітелію слизових оболонок та шкіри, а при його нестачі відбувається їх патологічне ороговіння — кера-тинізація, тому з'являються ксерофтальмія, кон'юнктивіт, ураження слизових оболонок слинних залоз, дихальних шляхів (бронхіт, бронхопневмонія), органів травлення (гастрит, ентерит), статевих органів (вагініт, ендометрит). Вітамін А бере участь в обміні речовин (білків, ліпідів, вуглеводів, макроелементів), імуногенезі, підвищує стійкість організму проти інфекцій та інвазій і фагоцитарну активність нейтрофілів, стимулює репродуктивну функцію самців та самок, сприяє нормальному розвитку плода, його формуванню. Зважаючи на такі багатогранні функції вітаміну А, визначення каротину та ретинолу є обов'язковим елементом диспансеризації сільськогосподарських тварин.
Вміст каротину в сироватці крові визначають за Карром-Прай-сом у модифікації Юдкіна, методом В. Ф. Коромилова і Л. О. Куд-рявцевої. В останні роки у лабораторіях каротин і вітамін А визначають за методом Бессея у модифікації А. А. Анісової на спектрофотометрі або на колориметрах КФК-2, КФК-3 (Левченко В. І., Сахнюк В. В., Щуревич Г. О., 1993).
Рівень каротину в сироватці крові залежить від виду й віку тварин. У свиней, дрібної рогатої худоби, кролів, собак є лише залишки каротину, тому кількість їх за методом В. Ф. Коромис-
268
лова виявити неможливо. Виходячи з цього, визначення вмісту каротину у тварин даних видів діагностичного значення не має. Аналогічні результати аналізу сироватки крові у новонароджених телят (1—5 мкг/100 мл) і телят місячного віку (до 20 мкг/100 мл). В подальшому відбувається безперервне зростання рівня каротину до 250—600 мкг/100 мл у молодняка 2—6-місячного віку і 450— 2000 мкг/100 мл у корів і молодняка старше б-місячного віку. В пасовищний літньо-осінній період вміст каротину у сироватці крові корів повинен бути не меншим 900 мкг/100 мл.
Низький вміст каротину в сироватці крові корів — гіпокароти-немія (менше 450 мкг/100 мл) є показником недостатнього надходження провітаміну у складі кормів раціону, руйнування його антивітамінами в передшлунках і кишечнику, порушення засвоєння у тонких кишках при їх запаленні або патології печінки. Критичним рівнем для корів є вміст 300 мкг у 100 мл сироватки крові (А. А. Душейко, 1989).
Зважаючи на обмежену діагностичну інформативність визначення каротину,'в практиці ветеринарної медицини все чільнішого місця набуває визначення ретинолу в різних субстратах: сироватці крові, молозиві, печінці, жовтках яєць (див. главу XI). У здорових новонароджених телят у перший день життя концентрація вітаміну А в сироватці крові становить 4—12, у 7—10-денних — 9—15 мкг/100 мл, місячних—12,5—25 (Левченко В. І., Сах-нюк В. В.), у 6—12-місячного молодняка — 30—80 (Щуревич Г. О., 1986)), у корів в стійловий період — 25—80, літній — 40—150, в овець— 15—25; коней— 10—17 (Ємеліна Н. Т. з співавторами, 1970); у новонароджених поросят—10—40; поросят-сисунів до місячного віку—15—50, відлучених та свиней на відгодівлі — ЗО—70, свиноматок — 20—50 (за Самохіним В. Т. з співавторами).
Зниження рівня вітаміну А в сироватці крові — А-гіповітаміноз спостерігається при недостатньому надходженні каротину, порушенні перетворення його у вітамін А при хронічному запаленні слизової оболонки кишечника, нестачі в раціоні протеїну, кобальту, йоду, легкорозчинних цукрів, наявності в кормах антивітамінів (нітритів, хлоридів нафталіну та ін.). Хвороби печінки (гепатоз, гнійний гепатит) будь-якої етіології викликають розвиток ендогенного або вторинного А-гіповітамінозу (Щуревич Г. О., 1986), оскільки при них зменшується секреція жовчі, синтез і виділення жовчних кислот, які беруть участь в емульгації ретинолу, гідролітичному розщепленні його ефірів, солюбілізації продуктів гідролізу і транспорті їх до кишкових епітеліоцитів.
269

ВИЗНАЧЕННЯ ЗАГАЛЬНОГО КАЛЬЦІЮ У СИРОВАТЦІ КРОВІ
У тілі дорослих тварин міститься 1,2+1,5 % кальцію з розрахунку на свіжу тканину. Основна маса його (близько 90 %) знаходиться у кістковій тканині в складі кристалів гідроксиапатиту, решта — в позаклітинній рідині, в тому числі плазмі крові. Крім формування скелету, кальцій бере участь у процесах зсідання крові, забезпечує нормальну збудливість і скоротливість різних м'язів, посилює систолічне скорочення серця, підвищує тонус серцевого м'яза, захисні функції організму, тонус симпатичного відділу вегетативної нервової системи, знижує проникність кровоносних судин.
В організм кальцій надходить у складі рослинних кормів і мінеральних добавок. Під впливом шлункового соку основна частина його перетворюється у кальцію хлорид, який є основною формою для абсорбції в тонкому кишечнику. Безпосередній механізм перенесення іонів кальцію через ентероцити до цього часу не з'ясований. Доведена участь у цьому процесі кальційзв'язуючого білка, що синтезується ентероцитами під впливом біологічно активних метаболітів вітаміну D3—1,25- та 24,25-дегідроксихолекаль-циферолу. Важлива роль у абсорбції важкорозчинних солей кальцію належить жовчним кислотам — холевій та дезоксихолевій.
Із організму кальцій, виводиться через кишечник, у лактую-чих тварин — з молоком, незначна кількість — з сечею.
У сироватці крові кальцій міститься в основному в двох формах: колоїдній, зв'язаній з білками (близько 40 % від загального кальцію), яка є біологічно неактивною, і у дифундованій, що складається з іонізованого кальцію (55%) і кальцію, зв'язаного з органічними аніонами (5%). Для оцінки стану кальцієвого обміну в сироватці крові частіше визначають вміст загального кальцію, а рівень іонізованого розраховують за формулою Й. Тодо-рова.
Кількість загального кальцію у сироватці крові визначають за допомогою атомної абсорбційної спектрофотометрії (рис. 71), автоматичних біохімічних аналізаторів і комплекснометричними методами з трилоном Б та індикаторами флуорексоном (за Віче-Вим €. П. і КаракашоВим Л. В.) або мурексидом (за Луцьким Д. Я-). Вміст загального кальцію у сироватці крові наведено в таблиці 11. Вміст кальцію в сироватці крові птиці залежить від віку й періоду продуктивності. В сироватці крові курчат денного віку він становить 10—12 мг/100 мл (2,5—3 ммоль/л); індиченят, гусенят і каченят— 12—16 (3—4 ммоль/л), у несучок всіх видів у непродуктивний період — 9—12 мг/100 мл (2,25— З ммоль/л), а в період яйцекладки у курей до 40 (10), індичок, качок і гусок до 46 мг/100 мл (11,5 ммоль/л).
270

|
|
|
 Скачать 2.87 Mb.
Скачать 2.87 Mb.